Загорская В.В.
Институт экологической экспертизы и биоинформационных технологий.
Республика Башкортостан, 450571, Уфимский район, сан. Юматово, ул. Парковая, д. 36.
E-mail: Valeria76@mail.ru.
// Редкие и исчезающие виды животных и растений Республики Башкортостан. 2014. № 6. С. 25-27.
Данные орнитологического обследования орнитофауны Южно-Уральского региона, проведенного в 1974-1976 гг. В.Д. Ильичевым и В.Е. Фоминым (1988), не содержат сведений о ходулочнике. Впервые, о встрече с ним на территории Республики, упоминает В.А. Валуев (2001). Он отметил гнездование этого вида на территории Хайбуллинского района (неподалеку от Маканского водохранилища) в 2000 г.
На основании этих данных вид внесен Красную книгу Республики Башкортостан (2004), хотя на авторство В.А. Валуева ссылки не последовало. Поэтому ходулочник внесён автором очерка Красной книги как залётный вид. С начала XXI в. этот вид стал регулярно гнездится в Башкирии, осваивая территории Зауралья и Предуралья (Валуев, 2007). О.А.Торгашов (2002) встретил несколько гнездящихся пар в Мелеузовском районе республики.
В.А. Валуев (2005) указывает, что по сообщениям некоторых людей, этот кулик был встречен в Чишминском районе (устн. сообщение А.А. Крыгина), который наблюдал его в 2002 г.; ходулочника в Баймакском районе в окрестностях г. Сибай (А.Р. Ишбирдин); . в Кушнаренковском районе в 2003 г. (Л.А Едренкина ). В мае 2003 г. В.А. Валуев (2005) зарегистрировал гнезда четырех пар ходулочников. В 2006 г. выводок был отмечен в Чишминском районе Предуралья Башкирии, а в 2008 г. на фильтрационных прудах наблюдались одна взрослая и три молодые птицы, а в июне этого же года две молодые птицы зарегистрированы в окрестностях поселка Карламан Кармаскалинского района республики (Валуев, 2008).
Нами пара птиц была встречена на оз. Мулдаккуль Абзелиловского района республики 17 июля 2014 г.
Таким образом, статус ходулочника определяется, скорее, как чрезвычайно редкий, гнездящийся, но никак не «залетный» вид». Поэтому, правильнее было бы исключить ходулочника из списка залетных видов птиц, занесенных в Красную книгу Российской Федерации и отмеченных на территории республики Башкортостан (приложения 1 к Красной книге РБ) и включить этот вид в основной список животных, подлежащих охране на данной территории. Последнее необходимо в связи с единичными встречами этой птицы на территории республики, и по причине необходимости охраны основных мест обитания этого вида – малых водоемов, небрежное использование которых приводит к обеднению их биоразнообразия (Валуев, 2007а).
Литература:
Валуев В.А., Валуев К.В. Новые наблюдения редких птиц в Башкирии // Итоги биологических исследований. Вып. 6. 2000 г.: Сборник научных трудов // Изд-е Башкирск. ун-та. Уфа, 2001. С. 161-163.
Валуев В.А. Кулики (Limicoli) Башкортостана // Вестник Башкирского университета. 2005, № 2. С. 48-55.
Валуев В.А. Фауна куликов Башкортостана и ее изменения // Тезисы докладов VII Международного совещания «Достижения в изучении куликов Северной Евразии». 5-8 февраля 2007 г. Мичуринск, Мичуринский гос. пед. ун-т, 2007. С. 14-15.
Валуев В.А. Экологические проблемы малых водоёмов Башкортостана // Башкирский орнитологический вестник. Вып. 4. Уфа, РИЦ БашГУ, 2007а. С. 5-7.
Валуев В.А. К распространению ходулочника в Башкортостане // Материалы к распространению птиц на Урале, в Предуралье и Западной Сибири. Екатеринбург, изд-во Урал. ун-та, 2008. С. 18.
Ильичёв В.Д., Фомин В.Е. Орнитофауна и изменение среды (на примере Южно-Уральского региона) // Изд-во «Наука», 1988. 246 с.
Красная книга Республики Башкортостан. – Уфа. Башкортостан, 2004. 180 с.
Торгашов О.А. Гусеобразные и Ржанковые поймы реки Белой в черте города Мелеуза // Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. Екатеринбург: Изд-во «Академкнига», Екатеринбург, 2002. С. 254–258.
Хабибуллин В.Ф.
// Материалы по флоре и фауне Республики Башкортостан: Научный журнал. Вып. I. Уфа: РИЦ БашГУ, 2010. С.23-27.
Наличие эффективных методов обнаружения, учета и сбора животных зависит многое в эколого-фаунистических исследованиях.
Несмотря на то, что в настоящее время активно разрабатываются методы отлова беспозвоночных, в том числе природосберегающие (Цуриков, Цуриков, 2001), для ряда групп насекомых нет эффективных методов учета и сбора. Отсутствие же отработанных методик сильно осложняет проведение исследований, в том числе количественного учета. Например, трудно квантифицировать насекомых – дендробинтов, и для учета сугубо древесных обитателей применяют мало подходящий для этих целей метод кошения энтомологическим сачком (Биньковская, 2004).
Именно отработанная методика во многом способствует большому количеству работ, например, по жужелицам: с помощью ловушек Барбера можно быстро и надежно осуществить не только качественное изучение видового состава, но и получить количественные данных об обилии, численности, доминировании и т.п.
Традиционная сфера применения ловушек Барбера – сбор герпетобионтных насекомых (активно перемещающихся по поверхности грунта), преимущественно жужелиц (Палий, 1970; Фасулати, 1971 и др.).
Возможно ли расширить сферу применения таких эффективных методов учета и сбора, как ловушки Барбера, и на другие семейства насекомых?
Мы исходим из предположения, что подобные успешные технологии учета насекомых должны расширять сферу применения: не ограничиваясь только жужелицами, но оценить эффективность метода для других групп беспозвоночных; опыт такой апробации и излагается в данной статье.
МАТЕРИАЛЫ И МЕТОДЫ
Материал собран в июле 2010 года в окрестностях д.Калиновка Давлекановского района Республики Башкортостан.
Мы использовали ловушки Барбера для сбора всех насекомых, доступных для данного типа ловушек.
В качестве ловушек Барбера мы использовали прозрачные пластиковые стаканчики объемом 200 мл; на дно наливалось немного 4%-го формалина. Ловушки закладывались линейной трансектой через каждые пять метров в трех биотопах: тополевая лесополоса, пойма р.Демы, степной участок. Осматривались ежедневно. Отработано 74 ловушко-суток.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЯ
Из интересных встреч отмечу наличие титтасписа и фронталиса.
Нами обнаружены представители пяти классов животных: паукообразных, многоножек, насекомых, моллюсков, млекопитающих (рис. 1). Среди насекомых отмечены представители пяти отрядов.
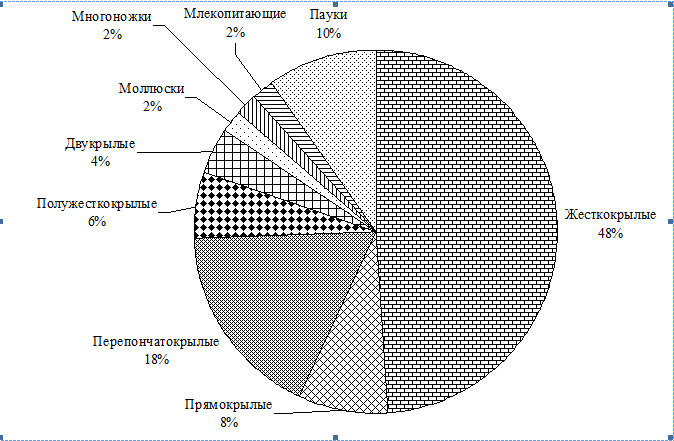
Рис. 1. Распределение отловленных ловушками Барбера видов животных по крупным таксонам.
Из рисунка 1 видно, что подавляющее число видов (93%) относятся к классу насекомые. Интересно, что в ловушки попадаются не только герпетобионты и хортобионты, но и типичные аэробионты – насекомые двукрылые и перепончатокрылые.
Небольшое количество особей некоторых видов попадается в очень больших количествах: это касается в первую очередь муравьев, пауков и некоторых видов жесткокрылых. Последних отловлено 403 особи, первых и вторых – несколько сотен. Без учета этих трех групп распределение особей остальных крупных таксонов показаны на рис. 2.
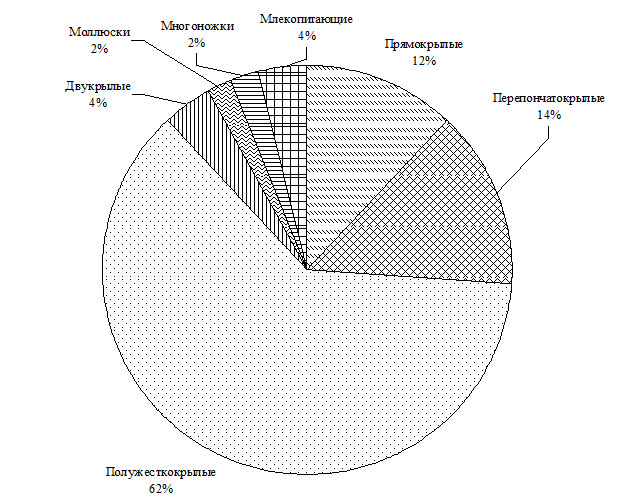
Рис. 2. Распределение отловленных ловушками Барбера животных по числу особей (без учета жуков, пауков, муравьев).
Из рисунка 2 видно, что после полужесткокрылых (62%) значительное количество особей относятся к перепончатокрылым (14%) и прямокрылым (12%).
Среди жуков отмечены представители 10 семейств (рис. 3).
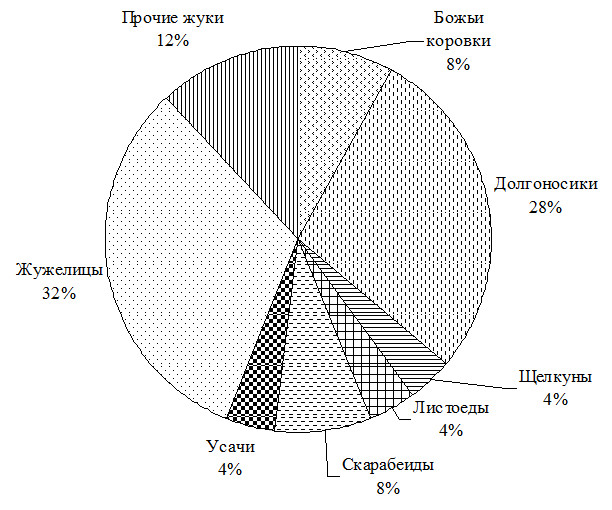
Рис. 3. Распределение отловленных ловушками Барбера видов по семействам жуков.
Показательно, что видовой состав долгоносикообразных жуков незначительно уступает жужелицам как по числу видов (7 против 8), так и по числу особей (60 против 80).
Показательны данные по использованию ловушек Барбера в широколиственных лесах заповедника «Калужские засеки» (Алексеев, Тиньгаева, 2002), где основными методами сбора были укосы, ручной сбор, сбор ловушками Барбера, разборка почвенных проб. Из общего числа обнаруженных видов (32) в ловушки попадались представители 17 видов (53,1%), в том числе семь видов (21,9%) известны только по единственному экземпляру, пойманному в ловушке Барбера: Stethorus punctillum, Scymnus (Pullus) auritus, Hyperaspis repensis, Сhilocorus bipustulatus, Exochomus quadripustulatus, E.flavipes, Semiadalia undecimnotata. Различными методами сбора, в том числе в ловушке Барбера поймано 10 видов (31,3%): Scymnus (Pullus) ferrugatus (два из 4-х экземпляров), Nephus redtenbacheri и Coccidula rufa (один из двух), Сhilocorus renipustulatus, Hippodamia septemmaculata, Tyttaspis sedecimpunctata (два из трех), Adalia bipunctata, Coccinella magnifica, Harmonia quadripunctata (один из трех), Halyzia sedecimguttata.
Именно ловушками Барбера удалось обнаружить несколько редких видов кокцинеллид, в том числе виды, известные только из сборов ловушками Барбера и никакими другими.
Обратим внимание но то, что при долговременной работе ловушек в них попадают и редкие виды, и ценность ловушек увеличивается для фаунистических исследований.
Таким образом, можно рекомендовать использование ловушек Барбера в качестве дополнительного средства при изучении локальных фаун пауков, жуков (жужелиц, долгоносиков), муравьев и отчасти других групп перепончатокрылых.
Относительно других групп наземных беспозвоночных что-либо определенное можно сказать лишь при проведении долговременных исследований.
БЛАГОДАРНОСТИ
Благодарю директора охотохозяйства «Рада» В.В.Аброщенко и директора Зоомузея БашГУ В.А.Валуева за помощь в организации работ; магистров кафедры зоологии БашГУ А.Маслову и Э.Давлетгареева за помощь в сборе материала.
Литература
1. Алексеев С.К., Тиньгаева О.В. Божьи коровки (Coleoptera, Coccinellidae) широколиственных лесов заповедника «Калужские засеки» // Известия Калужского общества изучения природы местного края. Кн. 5. Калуга, 2002. – С.172-179.
2. Биньковская О.В. Жуки-кокцинеллиды лесных экосистем юга Среднерусской возвышенности. – Дис. … канд. биол. наук. – Воронеж, 2004. – 189 с.
3. Палий В.Ф. Методика изучения фауны и фенологии насекомых. – Воронеж, 1970. – 312 с.
4. Фасулати К.К. Полевые изучения наземных беспозвоночных. – М.: Высшая школа, 1971. – 424 с.
5. Цуриков М. Н., Цуриков С. Н. Природосберегающие методы исследования беспозвоночных животных в заповедниках России: Труды Ассоциации особо охраняемых природных территорий Центрального Черноземья России. Вып. 4. – Тула, 2001. – 130 с.
Хабибуллин В.Ф., Хабибуллин А.Ф.
// Редкие и исчезающие виды животных и растений Республики Башкортостан: Научный журнал. Вып. II. Уфа: РИЦ БашГУ, 2010. С.16-21.
Редкие виды жесткокрылых слабо изучены в Республике Башкортостан: в республи-канской Красной книге (2004) значатся пять видов жуков, четыре из которых «делегирова-ны» из старшей по рангу Красной книги Российской Федерации (2001).
Видовое богатство жуков-кокцинеллид (Coleoptera-Coccinellidae) в Башкортостане на-считывает, по нашим данным, 42 вида; однако сведения по редкости кокцинеллид в респуб-лике нет; соответственно не определен и природоохранный статус представителей этого се-мейства.
В Красной книге Республики Башкортостан (2004), равно как и в Красной книге со-седней Оренбургской области (1998) кокцинеллид нет. В Красную книгу Челябинской области (2005) включены три вида коровок, все с четвертым природоохранным статусом (малоизученные виды): Hyperaspis reppensis (Herbst, 1784), Hyperaspis (Oxynychus) erythrocephalus (Fabricius, 1787), Chilocorus renipustulatus (Scriba, 1790).
В данной работе мы приводим данные по редким видам жуков-кокцинеллид (Coleop-tera-Coccinellidae) открытых местообитаний Башкортостана.
МАТЕРИАЛ И МЕТОДЫ
Материал собран на территории пяти районов Республики Башкортостан; исследова-лись только «открытые» биотопы: луга, степи, безлесная пойма. Проанализированы данные из следующих точек:
Караидельский район (д. Павловка) пойма р.Уфы (июнь 2005 г.)
Мелеузовский (восточнее пос. Зирган) пойма р.Белой (июль 2009 г.)
Давлекановский (близ д.Константиновка) остепненные участки (июль 2010 г.)
Бурзянский (БГПЗ) степные участки, пойма (июль 2000 г.)
Кармаскалинский (окрестности д.Бишаул-Унгарово) открытые биотопы (июль 2008 г.).
Методы сбора: кошение энтомологическим сачком, стряхивание на полог, ручной сбор; в Давлекановском районе – дополнительно – ловушки Барбера. Учитывались только имаго. Систематика приводится по данным Зоологического института РАН (www.zin.ru).
Обилие видов приводится по шкале Е.В.Бутько, 2005, с изменениями: 1) многочис-ленный – с численным обилием более 15%; 2) обычный – от 5 до 15%; 3) редкий – менее 5 %.
Активность видов (оценка значимости вида в фауне) оценивалась по следующей шка-ле (модифицировано нами из Т.К.Туневой, С.Л. Есюнина, 2008) (табл. 1).
Таблица 1
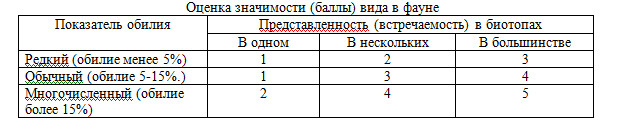
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В пяти обследованных участках обнаружено 18 видов кокцинеллид.
Видовой состав, обилие и активность видов представлены в таблице 2.
Таблица 2
Видовой состав, обилие, активность кокцинеллид открытых местообитаний Башкортостана
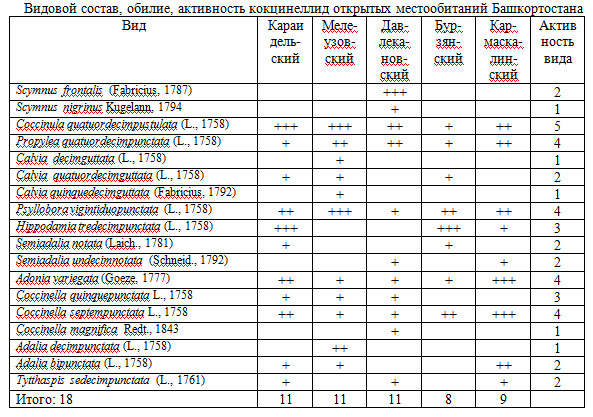
Выявленное видовое богатство кокцинеллид невелико; отсутствие вида в сборах может свидетельствовать о несовершенстве методики лова, сроков сбора, неудачности отдельных участков, продолжительности проведения работ.
Так, обнаружено только два вида сцимнин (род Scymnus s. lato). Как мы можем это объяснить? Считается, что сцимины приурочены в основном к древесной растительности (Дядечко, 1954), в открытых биотопах встречаются редко. С другой стороны, на степных участках Давлекановского района Башкортостана S.frontalis – обычный и, в момент наших наблюдений (июль 2010 г.) – самый многочисленный среди кокцинеллид вид; и явных при-чин для невыявления сопоставимых с ним по размеру гетероспецификов нет.
Что можно сказать о перспективах дальнейших фаунистических исследований? Обна-ружение видов из регионального пула – см. прогноз в нашей работе 2004 года (Хабибуллин и др., 2004). На тот момент список кокцинеллид насчитывал 36 видов, предполагалось наличие еще 29. Из этого прогнозного списка шесть видов уже добавлены, это: Stethorus punctillum Weise, 1891; Scymnus (s.str.) nigrinus Kugelann, 1794; Chilocorus bipustulatus (Linnaeus, 1758); Coccidula rufa (Herbst, 1783); Vibidia duodecimguttata (Poda, 1761); Harmonia quadripunctata (Pontoppidian, 1763); в том числе S.nigrinus – из обсуждаемых здесь сборов. «На очереди» – еще как минимум 23 вида.
Далее к вопросу о редкости кокцинеллид в регионе. Определение природоохранного статуса для насекомых имеет свою специфику (Дедюхин, 2008): считается, что не требуют принятия мер особой охраны редкие либо спорадично встречающиеся эврибионтные виды; виды, имеющие прогрессирующие ареалы; виды, характерные для пионерных стадий сукцессионого ряда (т.к. для поддержания данных экосистем необходим высокий уровень антропогенного воздействия, а не его ограничение).
К редким видам из собранных нами материалов (на основании оценки активности видов) мы можем отнести следующие пять: Scymnus nigrinus, Calvia decimguttata, Calvia quinquedecimguttata, Coccinella magnifica, Adalia decimpunctata. Из этого списка четыре вида считаются дендробионтами: оба вида кальвий, сцимнус и адалия. В лесных местообитаниях их встречаемость выше; в открытых же биотопах они оказались, скорее всего, случайно.
Таким образом, по обсуждаемым сборам, к редким видам открытых местообитаний Башкортостана можно отнести только коровку Coccinella magnifica.
Отметим необходимость дальнейших работ по уточнению природоохранного статуса выявлению редких видов насекомых в Башкортостане.
Благодарности
Благодарим Валиеву З.Р. за материалы по Кармаскалинскому району, администрацию БГПЗ, директору охотохозяйства «Рада» Аброщенко В.В – за помощь в организации полевых работ.
Литература
1. Бутько Е.В. Экология и внутривидовая изменчивость кокцинеллид (Coleoptera: Coccinellidae) в Восточном Забайкалье. – Дис. … канд. биол. наук. – Улан-Удэ, 2005б. – 187 с.
2. Дедюхин С.В. Рекомендуемые принципы охраны природного разнообразия насеко-мых на региональном уровне // Принципы и способы сохранения биоразнообразия: материалы III Всероссийской научной конференции / Мар. гос. ун-т. – Йошкар-Ола; Пущино, 2008. – С.136-137.
3. Дядечко Н.П. Кокцинеллиды Украинской ССР. – Киев: Изд-во АН УССР, 1954. – 153 с.
4. Красная книга Республики Башкортостан. Т.3. Животные / Под ред. М.Г. Баянова. – Уфа: Изд-во Башкортостан, 2004. – 180 с.
5. Красная книга Российской Федерации. Т. 1: Животные. – М., Астрель, 2001. – 863 с.
6. Тунева Т.К., Есюнин С.Л. Разнообразие фауны и географическое распределение пау-ков-гнафозид (Aranei, Gnaphosidae) Урала // Зоологический журнал, 2008. – Т.87. – №7. – С.779-789.
7. Хабибуллин В.Ф., Степанова Р.К., Хабибуллин А.Ф. Жуки-коровки (Coleoptera: Coccinellidae) Республики Башкортостан. Учебное пособие. – Уфа: Изд-во БашГУ, 2004. – 105 с.
Загорская В.В.
E-mail: valeria76@mail.ru
// Башкирский орнитологический вестник. 2012. № 10. С. 9-11.
Исследования динамики обилия сизого голубя и домового воробья в г. Уфе проводилось нами с января по октябрь 2012 г. Учеты проводились по методике Ю.С. Равкина (1967) в массивах старых многоэтажных застроек. Данный массив характеризуется наличием 5-этажных зданий с чердачными помещениями, построенных в 1950-70-е гг. Согласно градации, предложенной А.П. Кузякиным (1962), сизый голубь и домовый воробей относятся к весьма многочисленным видам в г. Уфе.
Обилие этих видов показано на графиках, где по оси х отмечены периоды: I, II, III и т.д. – месяцы; 1 и 2 – первая и вторая половина месяца соответственно.
Численность сизого голубя в 2012 г. колеблется в пределах от 69,57 ос./км² в мае до 256,52 ос./км² в январе; на протяжении всего года она непостоянна и имеет достаточно выраженную пульсацию (рис. 1). Аналогичная динамика обилия сизого голубя в зимний период по данным В.А. Валуева (Валуев, 2008) прослеживалась в г. Уфе в 1990 и 2002 гг.
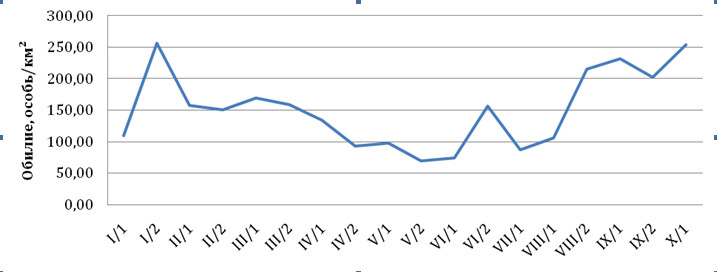
Рис. 1. Динамика обилия сизого голубя в 2012 г.
В 1990 г. максимальная численность сизого голубя приходилась на вторую половину января, в 2002 г. – на первую половину февраля. К весне численность его начинала снижаться. Такую же картину мы наблюдали и в 2012 г. Коэффициент перемещения «Km» (Валуев, 2007) сизого голубя в 2012 г. (0,97) практически равен таковому в прошлые периоды 1990 и 2002 гг. (Валуев, 2008), что указывает на стабильность популяции (табл. 1).
Наибольшая численность домового воробья в 2012 г., как и сизого голубя, приходится на зимние месяцы, а уже к марту снижается достаточно существенно. Уровень обилия популяции, установившийся в марте, незначительно колеблется в течение всего оставшегося года (рис. 2). В зимний период 1990 и 2002 гг. аналогичную картину изменения численности домового воробья в г. Уфе наблюдал В.А. Валуев (Валуев, 2008). Коэффициент перемещения «Km» домового воробья в 2012 г. составил 1,2. Его увеличение по сравнению с прошлыми периодами (табл. 1) указывает на начинающую дестабилизацию популяции домового воробья в городских условиях.
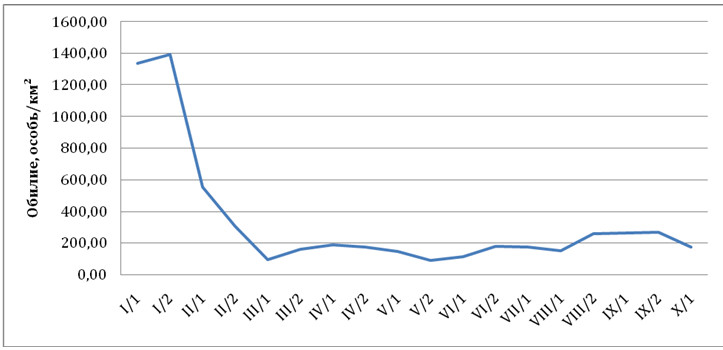
Рис. 2. Динамика обилия домового воробья в 2012 г.
Таблица 1.
Коэффициент перемещения «Km» для сизого голубя и домового воробья на примере г. Уфы
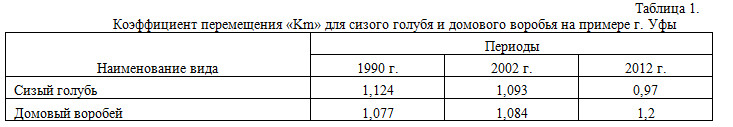
В 1990 г. максимальное обилие сизого голубя в январе составляло 1337 особи/км², в 2002 г. – 680,33 особи/км², в 2012 г. – 256,52 особи/км². Обилие домового воробья в январе 1990 г. достигало 2318 особей/км², в 2002 г. – 1168,83 особи/км², а в 2012 г. – 1391,3 особей/км². Таким образом, численность этих видов к 2012 г. сократилась соответственно в 5,2 и 2,7 раза, по сравнению с годами предыдущих исследований.
Данные по обилию показывают, что численность домового воробья в 2012 г. остаётся практически такой же, как и в 2002 г., в то время как коэффициент перемещения «Km» указывает на дестабилизацию вида.
Литература:
Валуев В.А. Экология птиц Башкортостана (1811-2008). Уфа, Гилем, 2008. 712 с.
Валуев В.А. Коэффициент перемещения вида и коэффициент стабилизации вида – индикаторы экологического состояния среды обитания популяции // Вестник охотоведения. Т. 4. № 2. М., 2007. С. 205-206.
Кузякин А.П. Зоогеография СССР // Учён. зап. Моск. обл. пед. ин-та им. Н.К. Крупской. Т. 109. М., 1962. С. 3-182.
Равкин Ю.С. К методике учёта птиц в лесных ландшафтах // Природа очагов клещевого энцефалита на Алтае. Новосибирск, Наука, 1967. С. 66-75.
Загорская В. В.
E-mail: valeria76@mail.ru
// Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири: Региональный авиафаунистический журнал. Вып. 17. 2012 г. Издательство Уральского университета. С.67-69.
Несомненно, орнитофауну городов нельзя рассматривать в отрыве от окружающей среды, потому что существуют как пульсации популяций (Валуев, 2007), так и кочевки птиц между городскими ландшафтами и их окрестностями. Последние относительно г. Уфы изучал В. А. Валуев (2012). Несомненно, сезонные перемещения свойственны как фауне естественных, так и селитебных территорий (Валуев, 2007а; 2012). В настоящей работе мы сравниваем результаты исследований В. А. Валуева (2003; 2003а; 2003б; 2008) и полученные нами в 2012 г. Для анализа динамики зимней орнитофауны города использованы данные по численности пяти наиболее многочисленных видов: сизый голубь Columba livia, галка Corvus monedula, серая ворона C. cornix, большая синица Parus major, домовый воробей Passer domesticus. Учеты проводили по методике Ю. С. Равкина (1967). С ее же помощью в 1990 и 2002 гг. были получены данные В. А. Валуева, с которыми сравниваются результаты наших исследований. В итоге динамика зимней орнитофауны Уфы прослежена с 1990 по 2012 гг. Выявлено ощутимое снижение обилия птиц. Заметное его сокращение происходит в последнее время не только вне поселений человека (Валуев, 2009), но и в таких крупных городах как Уфа.
Таблица
Динамика обилия доминирующих видов Уфы
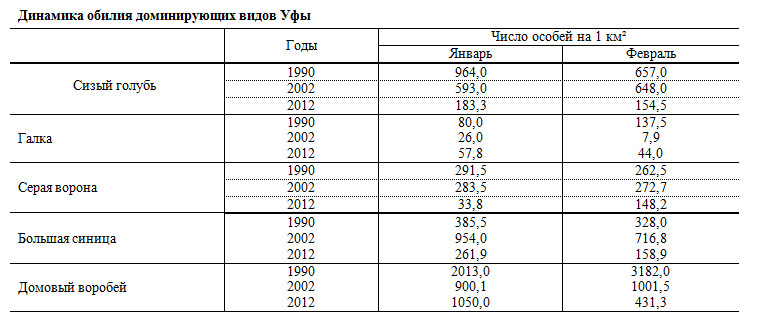
В таблице приведены данные по обилию доминирующих видов птиц Уфы за январь и февраль 1990, 2002 и 2012 гг. Результаты показывают, что за прошедшие 22 года численность сизого голубя сократилась в 5 раз, домового воробья – в 3,5, серой вороны – в 3, галки – в 2, большой синицы – в 1,7 раза. При этом за первый интервал (1990–2002 гг.) численность галки упала в 6,4 раза, домового воробья – в 2,7, сизого голубя – в 1,3; тогда как численность большой синицы выросла в 2,3 раза, серой вороны – осталась на прежнем уровне. За второй интервал (2002–2012 гг.) численность большой синицы снизилась в 4 раза, сизого голубя – в 3,7, серой вороны – в 3, домового воробья – в 1,2; численность галки выросла в 3 раза.
ЛИТЕРАТУРА
Валуев В. А. Динамика зимней авифауны г. Уфы. Препринт. Уфа, 2003. 12 с.
Валуев В. А. Зимние встречи птиц в Уфе в 2002 году // Итоги биологических исследований. 2001 г. вып. 7.: Сборник научных трудов. Уфа, 2003а. С. 51.
Валуев В. А. Птицы на зимних дорогах Башкирии // Вестник Башкирского университета. 2003б. № 3–4. С. 36–37.
Валуев В. А. Коэффициент перемещения вида и коэффициент стабилизации вида – индикаторы экологического состояния среды обитания популяции // Вестник охотоведения. 2007. Т. 4, № 2. С. 205–206.
Валуев В. А. Сезонные перемещения обычных и многочисленных видов орнитофауны Предуралья Башкортостана в весенне-осенний период // Вестник Оренбургского университета. Специальный выпуск. Оренбург, 2007а. Вып. 75. С. 56–61.
Валуев В. А. Экология птиц Башкортостана (1811–2008). Уфа, Гилем, 2008. 712 с.
Валуев В. А. Птицы семейств Oriolidae, Sturnidae,Corvidae, Bombycillidae, Cinclidae, Troglodytidae и Prunellidae, обитающие на территории Башкортостана // Вестник Башкирского университета. 2009. Т. 14, № 1. С. 76–79.
Валуев В. А. Сокращение численности птиц в Республике Башкортостан // Сборник научных трудов по материалам международной научно-практической конференции «Перспективные инновации в науке, образовании, производстве и транспорте 2011». Одесса, 2011. Т. 24. 77–79.
Валуев В. А. Взаимосвязь зимней орнитофауны города и его окрестностей (на примере доминирующих видов // Экология урбанизированных территорий. 2012. № 1. С. 74–79.
Равкин Ю. С. К методике учета птиц в лесных ландшафтах // Природа очагов клещевого энцефалита на Алтае. Новосибирск, 1967. С.66–75.
Валуев В.А., Полежанкина П.Г., Валуев Д.В.
// Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. Екатеринбург, издательство Уральского университета, 2007. С. 36.
Ранее чёрная ворона Corvus corone на территории Башкирии отмечалась в окрестностях г. Уфы П.П. Сушкиным (1897) и носила статус случайно залётного вида.
Нами зарегистрировано по одной особи чёрной вороны 19 и 22 апреля 1990 г. в окрестностях г. Нефтекамск и 29 мая 2004 г. – возле оз. Сатка в окрестностях д. Давлетово Абзелиловском р-на.
В окрестностях д. Баимово Абзелиловского р-на 29 апреля 2007 г. нами обнаружено гнездо, с которого слетела чёрная ворона. Гнездо располагалось в лесополосе на высоте 5-6 м. Кладка состояла из 5 яиц. В стенке лотка было несколько перьев совы. Слетевшая с гнезда птица не пыталась нас окрикивать и улетела. Второй взрослой птицы у гнезда не было. За двое суток в окрестностях д. Баимово мы несколько раз встречали одиночных чёрных ворон.
В.Ф. Хабибуллин
Применение понятия «зависимость» при изучении паразитизма позволяет напрямую измерить признак, по которому партнеры взаимодействуют (например, скорость, с которой паразиты потребляют ткани хозяина), т.е. что паразит на самом деле делает, а не последствия этого. Возможно изучение «зависимостей» по одному или нескольким признакам, для одной или нескольким парам партнеров.
В специальной литературе для определения понятия «паразитизм» привлекается, помимо прочих, признак «зависимость» (Cheng, 1986). Один из партнеров межвидового взаимодействия зависит – значит, испытывает необходимость, нуждается в другом партнере по какому-либо признаку. В данной работе мы рассмотрим возможность применения этого признака при изучении паразитизма.
Сначала проанализируем качественную зависимость по одному признаку, например, способность к синтезу какого-либо вещества. В исходной ситуации один из партнеров способен, а второй не способен к синтезу некоего вещества, необходимого каждому из них. При неспособности к самостоятельному синтезу второй партнер получает данное вещество от первого, т.е. зависит от него по этому признаку. Помимо крайних состояний (абсолютная зависимость или независимость), можно выявить промежуточные значения и выразить их количественно: один из партнеров зависит от другого по данному признаку на х% (=удовлетворяет потребность в данном веществе на х% за счет второго партнера). Изменяя уровень способности к самостоятельному синтезу вещества (т.е. степень зависимости по этому веществу от партнера), можно количественно отразить континуальность этой зависимости от 0 до 100%. На шкале показано только область положительных значений (0 – 100%). Область отрицательных значений будет относиться уже к зависимости другого партнера (его свойство) от изучаемого нами первого партнера, поэтому нами не рассматривается.
Далее можно рассмотреть зависимость партнеров по нескольким признакам, по нескольким партнерам. Заключительный шаг – анализ одновременной зависимости по нескольким признакам (признаки 1, 2, 3, …) и по нескольким партнерам (партнеры А, Б, В, …), качественно и количественно. После анализа всех позиций зависимости получим «спектр зависимости». Партнер А (акцептор) зависит от партнера Б (донор) (по признаку 1 – на x%, по признаку 2 – на n% и т.п.), от партнера В (по признаку 1 на y%, по признаку 2 на z% и т.п.), от партнера Г и т.д. Аналогично: партнер Б зависит… В итоге мы можем составить своего рода «паспорт» зависимостей по каждому партнеру. У каждого организма как субъекта межвидовых отношений существует свой набор требований к потенциальному партнеру. Этот набор может быть узким, если имеется потребность (зависимость) только в одном или нескольких партнерах по одной или нескольким функциям; но может быть и широким, если имеется потребность во многих партнерах по многим функциям.
Теоретически два и более разных партнера могут одновременно удовлетворять зависимость по одному и тому же признаку за счет друг друга, в соответствующих пропорциях. Взаимозависимость партнеров может быть, а может и не быть симметричной (когда каждый из партнеров в равной степени зависит от другого); как правило, один из партнеров «суммарно» более зависим, чем другой. Организм как бы предоставляет многофакторную – по числу признаков, формирующих зависимость – лицензию на партнерство: кто наиболее точно подходит под спектр требований, кто наиболее полно способен удовлетворить зависимость по всем признакам в нужном объеме – тот и принимается как партнер.
Проиллюстрируем изложенное на примере отношений «паразит-хозяин». Известно, что гельминты способны поглощать из тела хозяина витамины группы В: тиамин, рибофлавин, кобаламин и фолиевую кислоту (Леутская, 1991). Существует принципиальная возможность количественно оценить, по каким именно витаминам группы В и на сколько процентов зависит данный вид гельминта от данного вида хозяина; зависимость гельминтов других видов (по этой же функции – не/способность к синтезу витаминов группы В) от этого же вида хозяина; зависимость изученных видов гельминтов от других видов хозяев по рассматриваемому признаку. При более широком подходе считается, что при паразитизме происходит редукция собственных ферментативных систем паразита, при условии более выгодного – с энергетической точки зрения – использования ферментативных систем хозяина (Крылов, 1991). При таком подходе возможен количественный анализ зависимости по отдельным ферментам и целым ферментативным системам для конкретных пар «паразит-хозяин». Другой пример – взаимозависимость между эндосимбионтной бактерией Buchnera aphidicola и ее хозяином, гороховой тлей (Thomas et al., 2009). Примерно треть всех метаболических реакций бактерии обслуживает синтез необходимых хозяину аминокислот. Более того, метаболизм бактерии изменен таким образом, что для синтеза необходимых бактерии пуриновых соединений сначала должен быть синтезирован необходимый ее хозяину гистидин. В таком ракурсе хозяин (тля) становится еще более зависимым от своего эндосимбионта (бактерии), но и последний меняется, включая в свой метаболизм «избыточный» продукт, необходимый ее хозяину. Сравнение с метаболизмом эталонной Escherichia coli позволяет количественно оценить (в 27%) изменение метаболизма B.aphidicola и, следовательно, степень зависимости хозяина.
Достоинством применения признака «зависимость» представляется перенос акцента с оценки исхода межвидовых взаимодействий на оценку (в том числе количественную) конкретных механизмов, которые слагают эти взаимодействия. Например, при количественной оценке взаимодействий «паразит-хозяин» измеряют не косвенные изменения (снижение) приспособленности хозяев (Oliver et al., 2009), а напрямую признак, по которому партнеры взаимодействуют (например, скорость, с которой паразиты потребляют ткани хозяина), т.е. что паразит на самом деле делает, а не последствия этого (Poulin, Combes, 1999).
Список литературы
Крылов Ю.М. Молекулярные и генетические основы паразитизма у эукариот: эволюция отношений в системе «паразит-хозяин» // Эволюция паразитов: мат-лы I Всесоюз. симпозиума. – Тольятти: ИЭВБ АН СССР, 1991. – С. 163-167.
Леутская З.К. Биохимические адаптации в системе паразит-хозяин // Эволюция паразитов: мат-лы I Всесоюз. симпозиума. – Тольятти: ИЭВБ АН СССР, 1991. – С. 158-162.
Cheng T.C. General parasitology. General Parasitology, 2nd edition. Orlando: Academic Press College Division, 1986. 827 p.
Oliver T.H., Leather S.R., Cook J.M., 2009. Tolerance traits and the stability of mutualism // Oikos. V. 118. № 3. P. 346-352.
Poulin R., Combes C., 1999. The concept of virulence: interpretations and implications // Parasitology Today. V. 15. №. 12. P.474-475.
Thomas G.H., Zucker J., Macdonald S.J., Sorokin A., Goryanin I., Douglas A.E. A fragile metabolic network adapted for cooperation in the symbiotic bacterium Buchnera aphidicola // BMC Syst. Biol., 2009. – № 3. – P. 24.
PERSPECTIVES OF APPLICATION OF CHARACTER “DEPENDENCE” IN THE RESEARCH OF PARASITISM
V.F. Khabibullin
Bashkir State University, Validi str., 32, Ufa, 450074, Russia, herpetology@mail.ru
Summary
The character “dependence” can be applied as a useful criterion for study of host-parasite interactions. This character allows studying the relationships itself (i.e. the speed of host’s tissues consumed by parasite), not its consequences.
Хабибуллин В.Ф.
Традиции и перспективы развития зоопсихологии в России: Материалы Всерос. конф. с междунар. участием – Пенза, 2007. – С. 141-143.
В процессе преподавания дисциплин о поведении животных зачастую приходится балансировать между биологическим и психологическим компонентами.
Преподавание «психологического аспекта» поведения животных представлено гораздо лучше: курс «Зоопсихология и сравнительная психология» является базовым, обязательным для изучения для студентов факультетов психологии. Существует продуманная программа Н.Н.Мешковой, базовый учебник К.Э.Фабри и ряд толковых учебников и пособий других авторов; особенно ситуация в этом плане улучшилась в последние годы.
Биологическая составляющая преподавания этологических дисциплин разработана слабее. Науки о поведении животных в вузах не являются базовыми, существуют далеко не во всех вузах и представлены в виде спецкурсов под разными названиями (этология, поведение животных и пр.); нет единой программы и общепризнанных учебников; пере-водные издания содержат разнообразный материал и структурированы по-разному. Собственно «этология» как образовательная дисциплина хаотична, а содержание курса зачастую «несет печать» исследовательских интересов и предпочтений лектора.
Мы попытались синтезировать подходы и представить в рамках экспериментального спецкурса для биологов под компромиссным названием «Биологические и психологические основы поведения животных». Ниже приведены контуры разработанной программы, в которой отражены наиболее существенные, на наш взгляд, элементы как зоопсихологии, так и этологии.
Комплекс наук, изучающих поведение животных. Методы изучения поведения животных. Поведение как систематический признак. Формы индивидуального поведения. Формы группового поведения. Механизмы поведения. Регуляция поведения. Адаптивность поведения. Проблемы возникновения психики. Эволюция поведения и психики: основные этапы развития психики животных в процессе эволюции. Уровни развития психики. Развитие поведения и психики в онтогенезе: эмбриональном и пренатальном периодах. Инстинктивное поведение. Классификация инстинктов. Научение. Классификация видов научения. Высшие формы научения, проблема интеллекта животных. Орудийная деятельность, язык животных. Биологическая ограниченность поведения животных.
Данная программа не претендует на полноту и «окончательность» и открыта для конструктивных замечаний и предложений.
