Сен
17
Author Archives: admin
Сен
05
Загриев Ильдус Мустафович
ООО НПФ «Уральский камень Эко»
- Сизый голубь Сolumba livia в Башкирии (2015)
- К биологии воробьёв в частных застройках (2016)
- Адмирал Рyrameis atalanta в условиях Башкортостана и вопросы его учёта (2016)
- К динамике некоторых птиц в антропогенных местообитаниях Башкирии (на примере села Юматово) (2016)
- Рецензия на очерки П.Ю. Горбунова «Отряд чешуекрылые» Красной Книги Республики Башкортостан, 2014 (2017)
- К весенней авифауне лесопарка им. Калинина (Уфа) (2017)
- К сорокам Башкирии (2017)
Июл
20
Хирономиды: личинки комаров звонцов
Садчиков А.П.
Садчиков А.П. Культивирование водных и наземных беспозвоночных (принципы и методы). – М.: Изд-во МАКС Пресс, 2009. 272 с. ISBN 978-5-317-02931-9
Садчиков А.П. Биотехнология культивирования водных беспозвоночных. – М.: Изд-во МАКС Пресс, 2008. 160 с.
Массовое культивирование личинок насекомых (тип Arthropoda, класс Insecta) в качестве корма для рыб связано с большими трудностями, обусловленными особенностями их биологии и жизненного цикла. Хирономиды (у аквариумистов и рыбоводов – мотыль) относятся к роду Chironomus = Tendipes, отряда двукрылых (Diptera, семейство Chironomidae). Хирономиды (или же звонцы) на стадии личинки и куколки обитают в воде, а на стадии взрослого насекомого (имаго) ведут наземный образ жизни. В качестве объектов искусственного разведения служат Chironomus plumosus и Ch.thummi, относящиеся к числу широко распространенных форм, обладающих выносливостью к неблагоприятным факторам среды.
Как и вся группа двукрылых, хирономиды развиваются с полным метаморфозом, последовательно проходя стадию яйца, личинки, куколки и крылатого насекомого (имаго). Самая длительная стадия – личиночная.
Хирономиды населяют пресные водоемы, но встречаются и в солоноватых водах (озерах, прудах, лужах, каналах, бассейнах, бочках, опресненных заливах морей и т.д.).
Тело личинонок хирономид состоит из хорошо обособленной головы и удлиненного туловища, разделенного на 13 сегментов. Первые три сегмента образуют грудной отдел, 10 последующих – брюшной (или абдоминальный).
Покровы тела гладкие и представлены тонкой и прозрачной кутикулой. На переднем грудном сегменте личинки имеется одна пара ложных ножек, несущих хитиновые крючки. При помощи этих ножек личинки передвигаются по субстрату, а также строят чехлики. Тело заканчивается 13-м сегментом, где расположены две ложные ножки – задние ложные ножки (подталкиватели), служащие для передвижения личинки.
В естественных условиях личинки хирономид живут в мягком илу. У берегов водоемов, где грунт более плотный, личинки населяют его верхний илистый слой. Вдали от берега, по мере увеличения толщи мягкого ила, они могут проникать вглубь до 40 см.
Присутствие грубых частиц личинки переносят с трудом, особенно на ранних стадиях развития, что связано с характером их передвижения и питания. При наличии пищи они растут в условиях мягких грунтов значительно быстрее.
Температура воды или грунта влияет на жизненные процессы хирономид. Их существование возможно при относительно широком диапазоне температур. К низкой температуре менее чувствительны личинки, постепенно адаптированные к холоду и зимующие в водоемах при температуре воды, близкой к нулю. Такие личинки легко переносят даже замораживание. Вместе с тем летом личинки могут выдерживать прогрев воды до 35оС и более.
При наличии градиента температур личинки избирают зону с 17-18оС, т.е. температуру, при которой в естественной обстановке происходит их массовое окукливание, вылет имаго и откладывание яиц.
Температура среды влияет на число генераций хирономид в течение года. В небольших, хорошо прогреваемых водоемах жизненный цикл хирономид укорачивается, и до наступления осеннего похолодания они дают 2-3 и даже 4-5 генераций. В глубоких озерах, где температура более низкая и стабильная, у хирономид в течение года бывает всего одна-две генерации.
Поддерживая благоприятную температуру в закрытом помещении можно получать кладки хирономид непрерывно в течение всего года, без какого-либо снижения интенсивности развития и размножения их в осенние и зимние месяцы.
Места обитания личинок хирономид отличаются низким содержанием кислорода. В илах водоемов количество кислорода редко достигает 3 мг/л, часто оказывается близким к нулю.
Устойчивость хирономид к таким условиям связана с наличием в их крови гемоглобина. Содержание гемоглобина у личинок изменяется в пределах 0,1-0,25 мг/г сухой массы в зависимости от условий среды. В поверхностных слоях ила, где кислорода больше, количество гемоглобина составляет около 8% сухой массы тела, тогда как у личинок, обитающих на большей глубине, – 27%.
Находясь в среде с низким содержанием кислорода, личинки становятся темно-красными. Содержание гемоглобина при этом достигает 30 мг/г сухой массы. Благодаря этому хирономиды заселяют места малопригодные для жизни других животных. При полном отсутствии кислорода личинки хирономид впадают в состояние анабиоза.
Наиболее благоприятным для личинок является содержание кислорода в пределах 1,7-3,0 мг/л. В связи с этим при культивировании хирономид кислородному режиму должно уделяться большое внимание. Достаточное его содержание в среде (6-8 мг/л) легко поддерживается при небольшом слое ила (1,0-1,5 см) и уровне воды над ним не более 1-2 мм.
Личинки хирономид выдерживают широкий диапазон изменений концентрации водородных ионов (рН).
Отношение к свету у личинок хирономид изменяется с возрастом. На ранних стадиях развития у пелагических личинок обнаруживается положительной фототаксис. Положительная реакция на свет сохраняется и у личинок, осевших на дно, вплоть до наступления III личиночной стадии. С появлением в крови гемоглобина реакция на свет становится отрицательной. Взрослые личинки активно избегают светлых мест. Отрицательная реакция на свет исчезает у хирономид на стадии превращения куколки в комара. В этот момент куколка покидает чехлик и поднимается к поверхности воды.
Нормальный рост и развитие личинок хирономид протекает лишь при определенной плотности. В естественных условиях концентрация личинок в небольших временных водоемах достигает у крупных видов (Chironomus plumosus) 90 тыс.экз./м2, а у мелких (Ch.thummi) – 400 тыс.экз./м2. Наибольшая биомасса хирономид наблюдается осенью и в начале зимы. Весной, в связи с вылетом комаров и зимней гибелью личинок, биомасса наименьшая. Колебание численности хирономид связано не только с периодом вылета комаров, но и выеданием их рыбами и естественной смертностью. Отмирание личинок во многом связано с высокой их плотностью (а также накоплением в среде метаболитов, дефицитом кислорода).
При достаточно высокой плотности (свыше 5 тыс.экз./м2) личинки хирономид обитают в основном в поверхностном слое грунта (0-5 см), тогда как при небольшой плотности (200-400 тыс.экз./м2) наибольшее число особей концентрируются в слое ила 5-10 см и глубже.
Самцы хирономид (имаго) несколько крупнее самок и имеют длинные густо опушенные усики. Как правило, в популяции самцов на 10-15% больше, чем самок. Появляясь из куколки, самки способны откладывать яйца уже через 20-30 часов. Откладке яиц предшествует оплодотворение, совершающееся при роении комаров. Рой образуется утром и в сумерках на высоте 2-3 м и состоит в основном из самцов и небольшого количества самок, которые время от времени влетают в него с прибрежных кустов. Копулирующие пары падают в воду, и вскоре после оплодотворения самки начинают откладывать яйца. Роение комаров осуществляется при теплой безветренной погоде; роение прекращается при плохой погоде и наступлении осеннего похолодания. При искусственном разведении копуляция у Chironomus thummi прекращалась при понижении температуры до 10-12оС.
Кладка представляет собой удлиненный слизистый «мешок», на конце которого имеется удлиненный жгут. Самка сбрасывает кладку в воду, которая опускается на дно или с помощью тяжа закрепляется на растениях. В воде студенистая масса разбухает, и яйца оказываются заключенными в слизистый футляр. Количество яиц в кладке варьирует в зависимости от размера самок, чем они крупнее, тем больше откладывают яиц. Яйца имеют вытянутую форму и расположены в кладке перпендикулярно к длине тяжа. Длина яйца в среднем 375 мкм, ширина 150 мкм. Самки Chironomus thummi откладывают сразу около 900 яиц, а Chironomus plumosus – до 1200-1500 яиц.
Отложив яйца самки вскоре погибают. Общая продолжительность жизни самок не превышает пяти дней, самцов – и того меньше.
Срок эмбрионального развития яиц длится 2,5-6 суток. При температуре 20оС выклев первых личинок наблюдается через 59 часов с момента откладывания яиц. Массовый выход личинок происходит спустя 140 часов. С повышением температуры продолжительность развития яиц заметно сокращается, однако одновременно с этим увеличивается их гибель. В естественных условиях гибель яиц достигает 98%.
Выклюнувшиеся из яиц личинки Chironomus thummi 10-20 часов остаются в слизистой массе кладок; личинки Chironomus plumosus остаются там 2-5 дней. Покидая кладку, личинки переходят к пелагическому образу жизни, продолжающемуся 3-5 дней, после чего оседают на дно, где и происходит их дальнейшее развитие, вплоть до окукливания.
У новорожденных личинок гемоглобин отсутствует, из-за чего их тело бесцветно. На ранней стадии постэмбрионального развития личинки имеют жировые включения, уменьшающие их удельную массу.
Формирование и рост личинок идет в процессе линек, которых у Chironomus thummi три. При благоприятных условиях жизни первая линька наступает через 2-3 дня с момента выхода личинки из кладки. С первой линькой заканчивается первая личиночная стадия, в течение которой происходят наиболее существенные изменения в морфологии. Вторая личиночная стадия продолжается 5-6 дней. В начале этой стадии личинки оседают на дно и при помощи паутиновых желез и хитиновых крючьев ножек строят себе чехлики. Третьей линькой, происходящей через 7-8 дней после второй, заканчивается третья личиночная стадия. В течение четвертой стадии у личинки формируются органы будущего комара. Длительность ее – 10-12 дней, после чего личинка превращается в куколку. Окукливание личинки происходит в чехлике.
Продолжительность отдельных личиночных стадий и всего личиночного периода зависит главным образом от температуры и условий питания. В естественных водоемах при благоприятных условиях (нормальное питание, температура 19-20оС) весь период линек заканчивается через 19-20 дней.
При температуре выше 18оС срок развития личинок сокращается, но одновременно повышается их гибель, достигающая при 25-28оС – 22-75%. При температуре 10оС срок развития личинок удлиняется до 75-120 дней. В естественных водоемах личинки, не претерпевшие всех положенных им линек к моменту наступления осеннего похолодания, в течение зимы остаются на одной и той же стадии развития и только весной, с повышением температуры, полностью заканчивают метаморфоз.
В зависимости от условий жизни личинки хирономид имеют разные размеры; Chironomus plumosus перед окукливанием достигает длины 30 мм и массы 90 мг. Chironomus thummi имеет меньшие размеры: в возрасте 5 дней личинки весят 1,2 мг, в возрасте 10 дней – 3,5 мг. Перед окукливанием (возраст 15 дней) средняя масса личинки составляет около 7 мг. Размер окукливающихся личинок в значительной мере зависит от температуры. При повышении температуры длина и масса личинок снижаются.
Куколка отличается от личинки передней частью тела, сильно расширенной и несегментированной, в которой заключены голова, груди, ноги и крылья будущего комара. Стадия куколки продолжается у Chironomus thummi 2-3 дня. Непосредственно перед завершением метаморфоза куколка выходит из чехлика и поднимается к поверхности воды, где и происходит вылет комара. Процесс вылупления очень короткий и длится всего несколько секунд.
Общий срок развития хирономид от выхода из яйца до вылета имаго в зависимости от условий среды продолжается от трех недель до нескольких месяцев.
Питание личинок хирономид начинается с момента выхода из кладки и продолжается в течение всех личиночных стадий до момента окукливания. Куколки, а также комары не питаются. Их развитие идет за счет резервных веществ, накопленных личинкой и жировых отложений.
В период планктонного существования (I и II личиночная стадии) личинки питаются взвешенными в воде частицами детрита. Переходя к донному образу жизни, они строят при помощи выделений слюнных желез рыхлые чехлики – трубочки. Внутри чехлика находится сеточка – тонкая паутинная перегородка. Ею личинки отлавливаются пищевые частицы – детрит и различные одноклеточные организмы. Наряду с такой фильтрацией имеет место и непосредственное поедание поверхностного слоя ила, содержащего значительное количество бактерий и других организмов (простейшие, водоросли, животные). При искусственном кормлении личинок водорослями, обнаружено слабое усвоение протококковых водорослей. Наибольшую пищевую ценность для личинок хирономид имеет ил и дрожжевые клетки. Суточный рацион варьирует в пределах 80-660% массы тела.
При культивировании хирономид в качестве пищи используют дрожжи, рыбную муку, сухую тонко измельченную растительность. Качество корма в значительной степени определяет темп развития и выживание личинок.
При обильном кормлении рост и общая продолжительность развития личинок сокращаются, однако появляется опасность резкого снижения в среде кислорода, приводящего к повышенной гибели животных. Наиболее благоприятные условия создаются в случае внесения 5-6 мг корма (кормовые дрожжи) на одну личинку в течение всего срока ее развития. При такой норме продолжительность личиночного развития не превышает 14-16 дней, а выживаемость выше 70%.
Тело личинок хирономид содержит большое количество белков, углеводов и минеральных веществ. Белки имеют полноценный состав аминокислот. В зольном остатке содержится значительное количество фосфора и железа. Отличаются хирономиды и относительно большим количеством витаминов (А, каротиноиды, В1, В2).
Газообмен у личинок хирономид происходит через покровы тела, но в основном осуществляется анальными жабрам, постоянно заполненными кровью. Находясь в чехлике, личинки совершают волнообразные движения, способствующие поступлению свежей воды. Интенсивность дыхания зависит от индивидуальных размеров, температуры и содержания в среде кислорода. С увеличением размера тела личинок снижается относительная скорость их дыхания. Способность хирономид длительное время существовать в анаэробных условиях, связана с особенностями их обмена. Окислительные процессы идут за счет кислорода, образующегося при специфических тканевых реакциях.
***
Личинки хирономид (мотыль) является прекрасным кормом для большинства видов рыб, поэтому их заготавливают в природных водоемах. Наибольший интерес для кормления взрослых рыб имеют личинки комаров-звонцов Chironomus plumosus, размер которых достигает 20-25 мм. Однако добывают и более мелких личинок.
Массовый вылет комаров начинается со второй половины мая по июнь включительно. Наиболее благоприятные периоды для заготовки мотыля – ранняя весна (сразу же после схода льда) до вылета первой генерации. Второй период промысла следует начинать с середины августа, вплоть до зимы. Особенно интенсивно заготовка должна осуществляться теплой осенью, т.к. в этих температурных условиях возможен очередной вылет комаров.
Наиболее излюбленные биотопы обитания личинок комаров – прибрежные илы на границе барьера водной растительности на глубине 1,2-2,0 метра. Эти глубины не промерзают, поэтому личинки в массе скапливаются в этих местах. В таких биотопах биомасса мотыля может достигать несколько килограммов на 1 м2.
Для заготовки мотыля пользуются черпаком с более или менее длинной ручкой, либо ведром на веревке и специальными ситами (в простейшем случае – решетом). Черпаком со дна зачерпывают ил и помещают его небольшими порциями в решето. При помощи вращательных движений избавляются от мелких частиц ила, после чего в решете остается мотыль и крупные частицы (грунт, растения, моллюски и др.). Сита на некоторое время вынимают из воды, затем опять осторожно опускают в воду. При этом некоторые личинки всплывают и их собирают небольшим черпаком. Повторив это несколько раз, в сито набирают новую порцию ила для промывки.
Добытый таким способом мотыль имеет значительную примесь различного мусора. Для получения чистого мотыля его помещают в сито с отверстиями такой величины, чтобы он мог пролезть через них; сито ставится на таз с водой так, чтобы оно касалось дном воды. Мотыль через отверстия выползает в воду, и падает на дно таза. Затем его собирают сачком и помещают в холщовую тряпку. Этот метод можно использовать для отделения живого мотеля от погибшего.
Для хранения живого мотыля можно пользоваться разными способами. Наиболее простой – хранение во влажной холщовой тряпке или мешковине в нижней части холодильника; при этом мотыль надо располагать слоем не более 1 см. Слегка влажный мотыль можно поместить в плотно прикрытый сосуд. Эти способы дают возможность сохранить мотыль в течение двух недель.
Хорошо сохраняется мотыль при содержании в низких сосудах с ежедневно сменяемой водой. Особенно благоприятные результаты получаются при содержании его в мелком песке с водой. Промытый песок располагают слоем 1-2 см в низком сосуде, заливают водой на 1-2 мм выше его уровня и помещают мотыль. Живой мотыль быстро закапывается в песок. При необходимости песок пропускают через сито, через которое он проходит, а мотыль остается. Мотыль хранят в холодильнике при температуре 3-5оС. Большие партии мотыля сушат или замораживают.
Скармливают мотыль рыбам, помещая его в плавающие кормушки с отверстиями внизу, через которые мотыль медленно выползает в аквариум или бассейн. Мотыль, который зарывается в грунт чаще всего погибает и, разлагаясь, портит воду.
***
Для увеличения кормовой базы рыбоводных прудов используют метод привлечения насекомых на свет. Насекомые при использовании соответствующих ламп летят с расстояния 1 км и более. Привлеченные таким образом насекомые спариваются и откладывают в водоем яйца, другие падают в воду и становятся кормом для рыб. Кроме того, насекомых можно отлавливать специальными ловушками и скармливать их рыбам.
Для улучшения условий, обеспечивающих массовую кладку яиц комарами, в прудах оставляют небольшие участки невыкошенной растительности (камыша, рогоза, тростника и др.). Эти места являются местом роения комаров. Если же пруды не зарастают растениями, то используют пучки соломы или небольшие плавающие сетчатые деревянные рамы, на которые укладывают тонким слоем скошенную наземную растительность. Рамы с растительностью устанавливают в разных местах каждого пруда. Роение и спаривание комаров происходит на заре и в сумерках.
В качестве излучателя предлагается использовать прямые ртутные кварцевые лампы, лампы дневного света с энергией излучения в спектральной области 380-320 нм. Лампы устанавливают на расстоянии 25-30 см (не более 50 см) от поверхности воды. Лампы устанавливают на разделительных дамбах прудов около водной растительности или искусственных растительных субстратов, что необходимо самкам комаров для откладывания яиц. Привлечение комаров на свет увеличивает в 5-10 раз численность и биомассу личинок хирономид в прудах.
Лёт насекомых начинается при температуре не менее 15оС. Привлекаются на свет представители 11 отрядов насекомых: двукрылые, ручейники, поденки, жуки, клопы, бабочки, перепончатокрылые, равнокрылые, сетчатокрылые, таракановые, прямокрылые. На 1 м2 может прилететь более 3 тысяч поденок, 150 ручейников, 22 тысячи комаров разных видов, 240 бабочек и других насекомых, составляющие за одну ночь около 100 г/м2.
Устанавливают люминесцентные лампы на прудах из расчета 1 лампа на 1-1,5 га прудовой площади.
Благодаря применению этих простейших способов привлечения комаров к прудам можно значительно повысить кормовую базу и тем самым улучшить условия питания рыб. Так, общая масса насекомых, привлеченных на свет за сезон на площади 1 га, может достичь 1 тонны.
А.П.Садчиков, Московский государственный университет
Аннотация
Садчиков А.П. Культивирование водных и наземных беспозвоночных (принципы и методы). – М.: Издательство МАКС Пресс, 2009, 272 с.
В книге обобщены сведения по биологии и методам лабораторного и производственного культивирования некоторых водных и наземных организмов: почвенных олигохет, нематод, коловраток, инфузорий, дафний, хирономид, личинок мух, мучных червей и др. Эти беспозвоночные служат кормом для промышленно разводимых рыб и обитателей аквариумов, домашней птицы, многих экзотических животных (земноводных, пресмыкающихся).
Представлены сведения по выращиванию навозных червей, что позволяет получать их биомассу и ценный продукт – биогумус. Описаны биологические особенности разводимых видов, их распространение в природе, отношение к факторам среды, размножение, рост, развитие, питание, продукционные характеристики, химический состав тела и пищевая ценность. При описании методов культивирования сообщаются сведения об основных биотехнических и нормативных характеристиках культур (плотность, поддержание оптимальных условий, уход, режим кормления).
Рассмотрены вопросы использования сырьевых ресурсов пресноводных водоемов для их использования в пищу животным, птице, рыбам – прибрежные растения, яйца артемий, ракообразные, донные беспозвоночные, гаммарус, сапропель и др. Эти сведения необходимы для рационального использования природных водоемов.
Книга предназначена для гидробиологов, ихтиологов, рыбоводов, специалистов рыбного хозяйства и охраны природы. Кроме того, книга представляет интерес для огромной армии аквариумистов, любителей животных, для начинающих предпринимателей, которые захотят заниматься культивированием живых кормов.
Книга содержит рисунки культивируемых организмов, схемы производственных циклов, список литературы.
Табл. 10, Илл. 41, Библиогр: 43 назв.
Июл
16
О куликах бассейна р. Белая
Кутлиахметов А.Н.
Башкирский государственный педагогический университет им. М. Акмуллы, ул. Октябрьской революции, 3а, г. Уфа
E-mail: Azat56@yandex.ru
// Башкирский орнитологический вестник. 2016. № 18. С. 57-61.
О куликах р. Белая и её притоков информация очень скудна. За сто лет (с 1981 по 1982 гг.) данные о них есть лишь у П.П. Сушкина (1891), С.В. Кирикова (1952), В.Д. Ильичёва и В.Е. Фомина (1988). Следует отметить, что о куликах р. Уфы данные предоставил Е.В. Карев (Ильичёв, Фомин, 1988). Он зарегистрировал в окрестностях 9 видов – малого зуйка, чибиса, черныша, травника, поручейника, перевозчика, мородунку, бекаса и вальдшнепа. В.А. Валуев (2005, 2006, 2006а, 2007) указывал на гнездование в Башкирии 19 видов, из которых 6 – ходулочник, большой улит, мородунка, дупель и степная тиркушка, являются чрезвычайно редкими. Таким образом, на долю других гнездящихся представителей этого подотряда приходится лишь 13 видов. В репродуктивный период на горных притоках р. Белая и в широтном её течении также зарегистрировано 13 видов: малый зуёк, чибис, кулик-сорока, черныш, фифи, поручейник, перевозчик, круглоносый плавунчик, турухтан, кулик-воробей, белохвостый песочник, бекас и вальдшнеп. Из них три вида (круглоносый плавунчик, кулик-воробей, белохвостый песочник) являются пролётными (Валуев, Полежанкина, 2007). В послегнездовой период (Валуев, 2007) в этом регионе РБ отмечено лишь 4 гнездящихся вида (малый зуёк, черныш, фифи, перевозчик) и два пролётных (кулик-воробей и белохвостый песочник).
На фоне общего снижения численности птиц (Валуев, 2011) и, в том числе, куликов (Валуев, Кунсбаева, 2011), количество некоторых представителей этого подотряда (турухтан, кулик-сорока, ходулочник, большой улит и степная тиркушка) в первой пятилетке XXI в. увеличивалось (Валуев, 2007а, 2008). А у таких видов, как малый зуёк, травник, бекас, дупель, вальдшнеп, большой и средний кроншнепы продолжала сокращаться.
Однако, вопреки бытующему мнению, что человек является основным фактором в исчезновении животных, не беря во внимание химизацию, от которой действительно погибает огромное количество птиц (Алфёрова, Долотовская, 1967; Валуев, 2004), изменение ландшафтов в сторону их мозаичности положительно влияет на авифауну (Валуев, 2005а, 2008, 2009; Валуев, Едрёнкин, 2005; Валуев, Полежанкина, 2011; наши наблюдения).
Исходя из того, что кулики часто летят в ночное время суток, регистрация их в дневное время не является конечной инстанцией. Однако, оставленные на берегу их перья, помогают в уточнении видового состава (Валуев, 2007б; Валуев, Ильичёв, 2009; Валуев, Шарафутдинова, 2009).
По нашим наблюдениям в черте г. Уфы на правобережье р. Уфы обитают, как минимум, 3 вида куликов (малый зуёк, чибис и черныш). Гнёзда двух последних видов мы не находили (у зуйка найдено 2 кладки), но они постоянно во время репродуктивного периода наблюдались здесь нами. Интересно, что в пойме р. Уршак (приток р. Белая) почти такой же видовой состав куликов (Валуев, 2013). В Иглинском районе (граничащим с Уфимским) регистрировались 5 видов куликов – малый зуёк, галстучник, чибис, травник и черныш (Валуев, 2014). Это не опровергает данных наших исследований, т.к. пребывание галстучника (вида гнездящегося в тундре) является, по нашему убеждению, случайностью.
На пролёте отмечены поручейник и бекас. Хотя последний, безусловно, должен гнездится в пойме Белой.
При определении птиц пользовались определителями В.К. Рябицева (2001) и В.А. Валуева (2009а).
Литература:
Алфёрова М.А., Долотовская У.А. Ядохимикаты и фауна // Научная конф. по вопросу биологии, посвящ. 50-летию Вел. Окт. соц. револ. Тез. докл. Уфа, 1967. С. 65-67.
Валуев В.А. Последствия химизации // Вестник охотоведения. Москва, 2004, Т.1, № 2, С. 211-212.
Валуев В.А. Кулики (Limicoli) Башкортостана // Вестник Башкирского университета. 2005, № 2. С. 48-55.
Валуев В.А. К вопросу об антропогенном влиянии (на примере отрядов Columbiformes, Cuculiformes, Apodiformes, Cocariformes в Республике Башкортостан) // Объединение субъектов Российской Федерации и проблемы природопользования в Приенисейской Сибири: тезисы и материалы докладов межрег. научно-практ. конф., 11-13 апреля 2005 г. / Краснояр. гос. ун-т. – Красноярск, 2005а.- С. 183-186.
Валуев В.А. Эколого-фаунистическая характеристика куликов Башкортостана и проблема их охраны. Диссертация на соискание учёной степени канд. биол. наук. Уфа, 2006. 183 с.
Валуев В.А. Эколого-фаунистическая характеристика куликов Башкортостана и проблема их охраны. Автореферат на соискание ученой степени кандидата биологических наук. Уфа, Изд-во РИЦ БашГУ. 2006а. 22 с.
Валуев В.А. Экология куликов Башкортостана и аспекты их охраны. Уфа: Гилем. 2007. 156 с.
Валуев В.А. К послегнездовому периоду пойменного орнитокомплекса Южного Урала // Горные экосистемы и их компоненты: Тр. междунар. конф. Часть 1. М., т-во научных изданий КМК, 2007. С. 119-121.
Валуев В.А. Фауна куликов Башкортостана и её изменения // Достижения в изучении куликов Северной Евразии: тезисы докладов VII Международного совещания по вопросам изучения куликов, г. Мичуринск, 5-8 февраля 2007 г. / ред.: П.С. Томкович, А.О. Шубин, А.Ю. Околелов. – Мичуринск: МГПИ, 2007а. С. 14-15.
Валуев В.А. К графической идентификации птиц по перьям (ГИПП) // Тезисы докладов VII Международного совещания «Достижения в изучении куликов Северной Евразии». 5-8 февраля 2007 г. Мичуринск, Мичуринский гос. пед. ун-т, 2007б. С. 12-14.
Валуев В.А. К распространению ходулочника в Башкортостане // Материалы к распространению птиц на Урале, в Предуралье и Западной Сибири. Екатеринбург, изд-во Урал. ун-та, 2008. С. 18.
Валуев В.А. Экология птиц Башкортостана (1811-2008). Уфа: Гилем, 2008. 712 с.
Валуев В.А. Роль антропогенного влияния на птиц отряда Ржанкообразные в Башкортостане // Бюл. МОИП. Отд. биол. 2009. Т. 114, вып. 3, приложение 1, ч. 1. С. 137-139.
Валуев В.А. Полевой определитель птиц Европейской части России и Урала (кроме птиц отрядов Буревестникообразные и Воробьинообразные). Уфа: ДизайнПолиграфСервис, 2009а. 224 с.
Валуев В.А. Сокращение численности птиц в Республике Башкортостан // Сборник научных трудов по материалам международной научно-практической конференции “Перспективные инновации в науке, образовании, производстве и транспорте 2011”. Том 24. Медицина, ветеринария и фармацевтика, биология, сельское хозяйство. – Одесса: Черноморье, 2011. 77-79.
Валуев В.А. К орнитофауне поймы нижнего течения реки Уршак (Башкирия) // Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. 2013. Вып. 18. С. 11-16.
Валуев В.А. К гнездящимся видам птиц Иглинского района Башкирии // Сборник научных трудов SWorld. Вып. 3. Т. 36. Одесса: КУПРИЕНКО СВ, 2014. ЦИТ: 314-014. С. 12-17.
Валуев В.А., Едрёнкин В.А. К антропогенным воздействиям на авифауну // Материалы III Международной научно-практической конференции. 12-13 мая 2005 г. Астрахань: Издательский дом “Астраханский университет”, 2005. С. 80-82.
Валуев В.А., Ильичёв В.Д. К методике определения птиц, после их попадания под винт двигателя летательного аппарата // Башкирский орнитологический вестник: Сборник статей. Уфа: РИО БашГУ, вып. 6. 2009. С. 11-12.
Кунсбаева Г.Б., Валуев В.А. Падение численности травника Tringa totanus в Чишминском районе республики Башкортостан // Экология и природопользование: прикладные аспекты: материалы I Региональной научно-практической конференции студентов, аспирантов и молодых учёных. Уфа, изд-во БГПУ, 2011. С. 45.
Валуев В.А., Полежанкина П.Г. К орнитофауне горной части Южного Урала // Горные экосистемы и их компоненты: Тр. междунар. конф. Часть 1. М., т-во научных изданий КМК, 2007. С. 122-125.
Валуев В.А., Полежанкина П.Г. Роль антропогенного влияния на птиц отряда Ржанкообразные в Башкортостане // “Состояние среды обитания и фауна охотничьих животных России”. Материалы V Всероссийской научно-практической конференции “Состояние среды обитания и фауна охотничьих животных России”, Москва 17-18 февраля 2011 г. М., 2011. С. 62-66.
Валуев В.А., Шарафутдинова Т.А. Фезеринг и проблемы его развития на Южном Урале // Роль классических университетов в формировании инновационной среды регионов. Фундаментальное естественно-научное образование – генерация знаний на базе научных исследований: Материалы Международной научно-практической конференции. 2-5 декабря 2009 г. Т. II. Ч. 2. Уфа: РИЦ БашГУ, 2009. С. 45-48.
Ильичёв В.Д., Фомин В.Е. Орнитофауна и изменение среды (на примере Южно-Уральского региона). М.: Наука. 1988. 247 с.
Кириков С.В. Птицы и млекопитающие в условиях ландшафтов южной оконечности Урала. М.: Изд-во АН СССР, 1952. 411 с.
Рябицев В.К. Птицы Урала, Приуралья и Западной Сибири. Екатеринбург: Изд-во Урал. ун-та. 2001. 608
Сушкин П.П. Птицы Уфимской губернии // Мат. к познанию флоры и фауны Российской империи. М., Отд. зоол., 1897. Вып. 4. 331 с.
Июл
16
К охоте могильника Aquila heliaca в Краснокамском районе РБ
Подмарёв А.И.
Охотничье хозяйство ООО фирма РОМ.
Е-mail: zveroboy57@mail.ru
// Редкие и исчезающие виды животных и растений Республики Башкортостан. 2016. № 12. С.18-19.
До сих пор сведения о питании хищных птиц на территории Башкирии скудны. Башкирские орнитологи обычно ссылаются на данные Г.П. Дементьева (1951), хотя этот автор про питание хищных птиц на территории Башкирии не указывал.

Рис. 1. Могильник несущий ежа (фото автора).
О питании хищных птиц на территории республики имеется лишь несколько статей (Валуев, 2008, 2011, 2014, 2015-2015в). Из них только четыре сообщают о питании могильника. В отличие от мнения Г.П. Дементьева о том, что основным кормом этого орла являются суслики, В.А. Валуев и др. (2004) утверждают, что в Башкирии могильник питается преимущественно врановыми птицами.
Нами 4 июня 2016 года в 3 км. к западу от села Николо-Берёзовка сфотографирован могильник несущий в лапах ежа (рис. 1).
Литература:
Валуев В.А. Экология птиц Башкортостана (1811-2008). — Уфа: Гилем, 2008. — 712 с.
Валуев В.А. Охота перепелятника на территории Башкортостана // Башкирский орнитологический вестник: Сборник статей. Уфа: РИО БашГУ, вып. 9. 2011. С. 5.
Валуев В.А. К питанию кобчика Falco vespertinus в условиях Башкирии // Русский орнитологический журнал 2014, Том 23, Экспресс-выпуск 1018: 2023-2025.
Валуев В.А. К охоте некоторых дневных хищных птиц Башкортостана // Башкирский орнитологический вестник: сборник статей. Вып. 15 (сентябрь). – Уфа: РИЦ БашГУ, 2015. С. 3-10.
Валуев В.А. К питанию хищных птиц Башкирии // Башкирский орнитологический вестник: сборник статей. Вып. 14. – Уфа: РИЦ БашГУ, июнь 2015а. C. 3-5.
Валуев В.А. К питанию могильника Aquila heliaca на Южном Урале // Русский орнитологический журнал 2015б, Том 24, Экспресс-выпуск 1186: 3221-3224.
Валуев В.А. Могильник Aquila heliaca — могильник // Башкирский орнитологический вестник: сборник статей. Вып. 14. – Уфа: РИЦ БашГУ, июнь 2015в. C. 5-8.
Валуев В.А., Галиева Л.Ф., Яковлев А.Г. Питание могильника в условиях центральных районов РБ // Итоги биологических исследований 2004. Выпуск 8: Сборник научных трудов. -Уфа: РИО БашГУ, 2004. С. 78-79.
Дементьев Г.П. Отряд хищные птицы Accipitres или Falconiformes // Птицы Советского Союза. М.: Изд-во «Советская наука». 1951. Т.1. С. 70-341.
Июл
02
К рациону питания тетерева Lyrurus tetriх в Башкирии
УДК 598.261
Валуев К.В., Загорская В.В.
E-mail: Valeria76@mail.ru.
// Материалы по флоре и фауне Республики Башкортостан. 2014. № 5. С. 26-27.
На фоне общего снижения обилия боровой дичи в республике любые данные по питанию тетерева представляют несомненный интерес. Подробное исследование питания тетерева проводилось только С.В. Кириковым (1952). В конце XX и начале XXI вв. по питанию птиц отряда Курообразные опубликованы лишь две работы (Бурзянцев и др., 1996; Феоктистова и др., 2013). По последним данным инвентаризации численности и распространения тетерева на территории республики Башкортостан (Валуев, Загорская, 2014) основная численность тетерева сосредоточена в Предуралье республики, в горах же он чрезвычайно редок. Согласно последним исследованиям годовой динамики боровой дичи на территории республики (Валуев, Дурягина, 2012) обилие тетерева осенью в горах начинает снижаться, увеличиваясь к весне в Предуралье, что можно объяснить кочевками этих птиц в поисках корма, – птицы летят кормиться на культурные посевы. Считается, что летом этот вид питается в основном беспозвоночными, а осенью переходит в основном на растительную пищу. У одной из особей, добытых в осенний период в Благовещенском районе, в желудке были обнаружены почки и сережки березы, у другой особи, залетевшей в Уфу, – почки, листья и ягоды калины (Валуев, 2008).
Нами было исследовано содержимое желудков и зобов двух тетеревов, добытых в сентябре 2014 г. в Архангельском районе Башкирии. По результатам определения растительных остатков можно сделать интересное дополнение к рациону питания тетерева в осенний период. Он включает:
– гречиха посевная Fagópyrum esculentum;
– зверобой продырявленный Hypéricum perforátum;
– звездчатка злаковая Stellaria graminea;
– мятлик луговой Poa pratensis;
– ягоды брусники Vaccínium vítis-idaéa;
– клевер луговой Trifolium praténse;
– подмаренник душистый Galium odoratum;
– будра плющевидная Glechóma hederácea.
В составе кормовых остатков преобладают семена гречихи посевной.
Выражаем благодарность за помощь в определении растительных остатков д.б.н., проф. кафедры ботаники А.Р. Ишбирдину.
Литература:
Бурзянцев А.В., Едренкина Л.А., Абдрахимов Р.Ф. Питание глухаря в условиях Башкортостана // Вопросы экологии жив. Ю. Урала. Уфа, 1996. Вып. 7. С. 70-76.
Валуев В.А. Экология птиц Башкортостана (1811-2008). Уфа: Гилем. 2008. 712 с.
Валуев В.А., Дурягина В.В. Динамика боровой дичи в Предуралье и горах Башкирии по сезонам // Вестник охотоведения. Том 9, № 1. Январь – Июнь 2012. С. 25-28.
Валуев В.А., Загорская В.В. К распространению тетерева Lyrurus tetrix осенью в Предуралье и горах Башкирии // Современные проблемы эволюции и экологии. Сб. мат. международной конф. Ульяновск: УлГПУ, 2014. С. 279-282.
Кириков С.В. Птицы и млекопитающие в условиях ландшафтов южной оконечности Урала. М., 1952. 412 с.
Феоктистова Я.А., Загорская В.В., Валуев В.А., Книсс В. А. Особенности осеннего питания рябчика Tetrastes bonasia (Linnaeus, 1758) в Республике Башкортостан // Башкирский орнитологический вестник. Вып. 11. Уфа, РИЦ БашГУ, 2013. С. 24-29.
Июн
19
К динамике Motacillidae в Республике Башкортостан
Валуев В.А.
Учебно-научный музей БашГУ.
// Русский орнитологический журнал 2014, Том 23, Экспресс-выпуск 1040: 2663-2668.
Аннотация
Показана динамика птиц двух подсемейств: Anthinae и Motacillinae. Отмечено, что численность всех гнездящихся на территории республики видов неуклонно падает. Зарегистрирована замена на территории Зауралья Башкирии одного подвида жёлтой трясогузки Motacilla flava bema, другим – M.f. flava.
Summary
Shows the evolution of birds two subfamilies: Anthinae and Motacillinae. Noted that the number of breeding species in the republic has been steadily declining. Registered in the territory of replacement Zauralye Bashkiria one subspecies Yellow Wagtail Motacilla flava bema, others – M.f. flava.
МАТЕРИАЛ И МЕТОДИКА
Данные о динамике обилия птиц семейства Трясогузковые Motacillidae более, чем за 100 лет (с середины XIX до второй четверти XX веков) взяты из трудов Э.А. Эверсманна (2001) – проводил исследования в середине XIX в., П.П. Сушкина (1897) – проводил наблюдения в 1891 г., С.В. Кирикова (1952) – проводил наблюдения в первой половине XX в., В.Д. Ильичёва, В.Е. Фомина (1988) – наблюдения проводили в 1974-1976 гг.
Оценка показателя обилия, которое отражено в письменных свидетельствах за столь долгий период, затруднена по причине того, что у предыдущих исследователей не были заложены общие подходы к количественному учету. В связи с этим, мы сопоставляли их свидетельства об обилии птиц изучаемой группы, в которых находили косвенные или прямые данные по численности вида, с количественной характеристикой шкалы балльных оценок обилия птиц предложенной А. П. Кузякиным (1962).
Например, о численности птиц можно судить по таким показателям, как количество добытых экземпляров, временной промежуток между встречами отдельных особей, количество в стаях и т.п., отражённых в публикациях этих авторов (Валуев, 2006).
Примеры сопоставительных оценок обилия по оценкам названных авторов и наших расчетных показателей отражены в таблице 2.
Таблица 1.
Пример подведения значений обилия птиц под единый показатель.
Показатели обилия по А.П.Кузякину (1962) |
Оценки обилия по письменным свидетельствам авторов XIX-XX вв., на примере П.П. Сушкина (1897) |
Весьма многочисленный |
«..на больших токах, возможно, добыть в одну ночь около сотни дупелей» (стр. 17) |
Многочисленный |
«Я нашёл перевозчика очень обыкновенным…» (стр. 25) |
Обычный |
«…кулик-сорока составляет необходимую принадлежность всех рек…» (стр. 32) |
Редкий |
«…большой улит довольно редок, но я летом неоднократно встречал его…» (стр. 28) |
Очень редкий |
«…мородунка…довольно редка, хотя и гнездится здесь. … добыл летом два экземпляра…» (стр. 25) |
Чрезвычайно редкий |
«Мне лично чернозобик не встречался. Пименов сообщил мне, что изредка чернозобик встречается всё лето…»(стр. 21) |
Результаты оценки динамики обилия птиц семейства Трясогузковые отражены на рис. 1 и 2. Отрезки оси ординат соответствуют принадлежности вида к разряду «очень редкий» – (-20) – (-10); «редкий» – (-10) – (0); «обычный» – (0) – (10), «многочисленный» – (10) – (20).
Мы проводили наблюдения за птицами на территории РБ с 1981 г; маршрутные учёты проводились по методике Ю.С. Равкина (1967) с 1989 г. При анализе общего обилия применялся понижающий коэффициент «ПК» (Валуев, 2004, 2006а, 2012). Русские и латинские названия, а также порядок следования видов приводится в соответствии со «Списком птиц Российской Федерации» (Коблик и др., 2006).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Анализ динамики обилия подсемейства Anthinae показал, что из четырёх видов коньков зарегистрированных на территории Республики Башкортостан луговой конёк Anthus pratensis на протяжении всего времени был чрезвычайно редким видом. Из предыдущих исследователей он был отмечен только П.П. Сушкиным как довольно редкий вид. Нами он встретился лишь дважды – 20 октября 1988 г. в Нуримановском районе (Валуев, 1989) и 20 сентября 2003 г. в Кугарчинском (Валуев, 2008). Чучело добытого в 1988 г. экземпляра хранится в зоомузее БашГУ.
Краснозобый конёк A. cervinus, будучи обыкновенен в XIX столетии, уже в начале XX в. регистрируется так редко, что С.В. Кириков и В.Е. Фомин наблюдали их лишь по одному разу – первый в горах, второй – в Зауралье республики. Мы регистрировали небольшие стайки этих птиц в Предуралье на весеннем пролёте в 1983 и 1986 гг. в Кармаскалинском районе. Две особи экспонируются в зоомузее БашГУ.

Рисунок 1. Динамика обилия коньков на территории РБ.
Полевой конёк A. campestris, чей основной ареал лежит южнее Башкирии, на территории республики в XIX в. практически не наблюдался. Как многочисленный вид он отмечен только С.В. Кириковым в начале XX в.; причём такое его обилие наблюдалось им лишь в Зауральском регионе республики и южной её оконечности. В.Е. Фомин регистрировал этот вид в 1974-75 гг., когда проводил учёты по Зауралью и югу Башкирии. По данным его экспедиции полевой конёк гнездился до 54°20́ с.ш. в Предуралье и в Зауралье республики. В начале XXI столетия этот вид был зарегистрирован лишь дважды: в 2004 г. в национальном парке «Башкирия» его наблюдал О.А. Торгашов (2004) и 6 июня 2006 г. на южной оконечности республики пару птиц встретила П.Г. Полежанкина (2006).
Нами полевой конёк был встречен только один раз – во время совместной экскурсии с немецкими коллегами в начале лета 1986 г. (Валуев,1989а) в окрестностях г. Стерлитамак. Птица совершала токовые полёты, поэтому можно предполагать, что это самая северная точка его гнездования в республике.
Лесной конёк Anthus trivialis считался многочисленным на протяжении последних двух веков. Но в конце XX столетия заметно наметился спад обилия этого вида. Причём тенденция к сокращению численности наблюдается из года в год (рис. 1); и в первых годах XXI в. он попадает в категорию «обыкновенный вид».
Анализ динамики видов подсемейства Motacillinae также указывает на имеющиеся колебания их численности; причём у некоторых видов довольно значительных. Особенно резко они проявляются у желтолобой Motacilla lutea и желтоголовой M. citreola трясогузок (рис. 2).
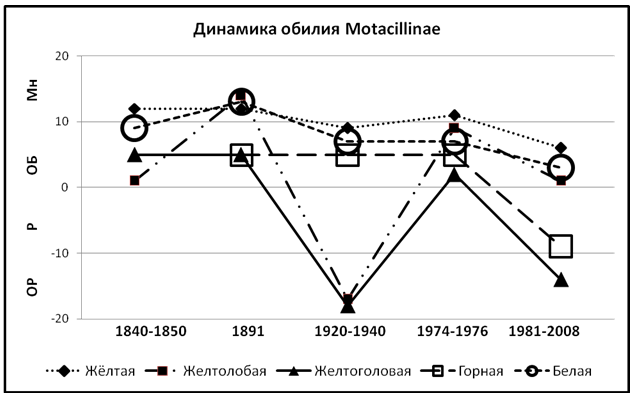
Рисунок 2. Динамика обилия трясогузок на территории РБ.
Динамика численности жёлтой Motacilla flava и белой Motacilla alba трясогузок, особенно если учесть погрешности учётов и их трактовки, довольно схожи. Большее обилие жёлтой трясогузки в последнее время, чем у белой, показанное на рис. 2, несмотря на то, что первая практически отсутствует на огромной территории северо-востока республики (Валуев, 2003), объясняется тем, что учёты проводились нами в естественных условиях, а большое количество белой трясогузки в Башкирии тяготеет к антропогенным, в том числе и к таким как очистные сооружения сахарных заводов или рыборазводные пруды (Валуев, 2004а).
Причём количество жёлтой трясогузки в таких селитебных районах как Уфимский, снижается и на протяжении гнездового периода, тогда как обилие белой трясогузки, сохраняется на одном уровне (Валуев, 2013). Здесь следует отметить, что практически полное отсутствие жёлтой трясогузки на северо-востоке республики (Валуев, 2004б) и не принятие ею селитебных районов компенсируется её преобладанием в несколько раз над M. alba на соседних нетронутых застройками территориях (Валуев, 2010, 2011а; Валуев, Едрёнкин, 2005) и многочисленностью в степях Зауралья республики (Валуев, 2010а). Также нужно указать, что в сезонной динамике всех видов трясогузок, зарегистрированных на территории Башкирии, только динамика обилия белой трясогузки отличается от других видов – ее обилие в репродуктивный период меньше, чем во время весенних и осенних миграций (Валуев, 2007). При поднятии птенцов на крыло, “желтые” откочевывают за пределы республики, а белая трясогузка остается на гнездовых участках до середины осени.
Это предположение подтверждают и учёты в горной области республики (Валуев, 2007а), где обилие жёлтой трясогузки в послегнездовой период в 18 раз меньше, чем у белой (0,5 и 8,9 особей/км² соответственно). В то время как в гнездовой период численность белой трясогузки в этом регионе республики превышала таковую жёлтой лишь в 2 раза (Баянов и др., 2005).
На территории Республики Башкортостан гнездятся только два подвида жёлтой трясогузки M.f. flava и M.f. bema. M.f. plexa была зарегистрирована на территории Башкирии только один раз – на пролёте весной 1983 г. в Кармаскалинском районе (Валуев, 1984).
Про состояние горной трясогузки Motacilla cinerea Э.А. Эверсманн не указывает, но предыдущими исследователями на протяжении последних 100 лет, с конца XIX в., она считалась обычным видом. Однако с конца XX в. численность её, как и у других видов трясогузок стала неуклонно падать и в 2006 г. составляла 0,02 особи/км² (Валуев, 2006б).
ЗАКЛЮЧЕНИЕ
Сокращение видов семейства Motacillidae одной кормовой базой объяснять не приходится. Такое снижение происходит у большинства видов населяющих республику (Валуев, 2011). Причём у жёлтой трясогузки отмечено замещение одного подвида другим. Например, M.f. beema, составляющая подавляющее большинство в Зауралье Башкирии с 2000 по 2005 гг., в 2006 г. уступила «пальму первенства» M.f. flava (Валуев, Артемьев, 2006). А это изменением лишь кормовой базы объяснить не представляется возможным. По всей видимости, происходят глобальные негативные изменения, связанные с отравлением окружающей среды, что сказывается как и на самой на продуктивности организма (Валуев, 2009), так и на нарушении его эндокринной системы (Fox at al, 1978).
ЛИТЕРАТУРА:
Баянов М.Г., Валуев В.А., Юмагужин Ф.Г. (2005): Птицы долины реки Белой в заповеднике «Шульган-Таш». Современные тенденции в биологических науках. Материалы Всероссийской научно-практич. конф. 15-16 сентября 2005 г. Бирск. 161-167.
Валуев В.А. (1984): Встречи редких птиц в Башкирии. – Вопросы экологии животных Южного Урала. Вып. 2. Уфа. 71-75.
Валуев В.А. (1989): К орнитофауне БАССР. – Распространение и фауна птиц Урала. Свердловск. 29-30.
Валуев В.А. (1989а): Некоторые результаты изучения редких видов птиц Башкирии. – Всес. совещ. по проблеме кадастра и учёта жив. мира. Уфа. Ч. 3. 36-37.
Валуев В.А. (2003): К распространению трясогузок в Башкирии. – Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. Екатеринбург, изд-во Уральского университета. 71-73.
Валуев В.А. (2004): Экстраполяционный коэффициент как дополнение к учёту численности по методике Ю.С. Равкина (1967) для территорий со значительной ландшафтной дифференциацией. – Вестник охотоведения. 1 (3): 291-293.
Валуев В.А. (2004а): К фауне птиц рыборазводных прудов Фёдоровского района Башкортостана. Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. Екатеринбург, изд-во Уральского ун-та. 44-49.
Валуев В.А. (2004б): Птицы северо-восточной области Башкортостана. Орнитологический вестник Башкортостана. Уфа, РИО БашГУ. Вып. 1. 2-9.
Валуев В.А. (2006). Эколого-фаунистическая характеристика куликов Башкортостана и проблема их охраны. Диссертация на соискание ученой степени кандидата биологических наук. Уфа, 2006. 183 с.
Валуев В.А. (2006а). О необходимости использования экстраполяционного (или понижающего) коэффициента. – Проблемы Красных книг регионов России. Материалы межрегион. науч.-практ. конф. 30 ноября – 1 декабря 2006 г. Пермь, Пермский ун-т. 234-236.
Валуев В.А. (2006б). К семействам Ласточковые, Жаворонковые, Трясогузковые и Сорокопутовые Башкортостана. Вестник Башкирского университета. Уфа. № 1. 48-54.
Валуев В.А. (2007): Сезонные перемещения обычных и многочисленных видов орнитофауны Предуралья Башкортостана в весенне-летний период. Вестник Оренбургского университета. Оренбург. Вып. 75. 56-61.
Валуев В.А. (2007а): К послегнездовому периоду пойменного орнитокомплекса Южного Урала. Горные экосистемы и их компоненты: Тр. междунар. конференции. Часть 1. М., Т-во научных изданий КМК. 119-121.
Валуев В.А. (2008): Экология птиц Башкортостана. Уфа: Гилем. 1-712.
Валуев В.А. (2009): Неудавшееся гнездование обыкновенного соловья. Башкирский орнитологический вестник. Вып. 7. Уфа, РИО БашГУ. 3-4.
Валуев В.А. (2010). Орнитофауна озера Асли-Куль. Материалы по флоре и фауне Республики Башкортостан: Вып. I. Уфа, РИЦ БашГУ. 35-38.
Валуев В.А. (2010а). Орнитофауна поймы Таналыка в среднем и нижнем течении. Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. Екатеринбург, изд-во Уральского университета. 34-35.
Валуев В.А. (2011). Сокращение численности птиц в Республике Башкортостан // Сборник научных трудов по материалам международной научно-практической конференции «Перспективные инновациии в науке, образовании, производстве и транспорте 2011». Том 24. Медицина, ветеринария и фармацевтика, биология, сельское хозяйство. Одесса, Черноморье, С. 77-79.
Валуев В.А. (2011а). Орнитофауна озера Каряжное и его окрестностей (Чишминский район Республики Башкортостан). Башкирский орнитологический вестник. Вып. 9. Уфа, РИЦ БашГУ. 3-4.
Валуев В.А. (2012): «За» и «против» «понижающего» коэффициента. – Сборник научных трудов SWorld. Материалы международной научно-практической конференции «Научные исследования и их практическое применение. Современное состояние и пути развития. 2012». Одесса: КУПРИЕНКО. Цит. 312-103. 31 (3): 36-43.
Валуев В.А. (2013): Сезонная динамика обилия трясогузок Уфимского района Башкирии в 2013 г. Сборник научных трудов SWorld. Материалы международной научно-практической конференции “Научные исследования и их практическое применение. Современное состояние и пути развития ‘2013”. Выпуск 3. Том 44. Одесса: КУПРИЕНКО. Цит: 313-0300. 51-54.
Валуев В.А., Артемьев А.И. (2006): К распространению трясогузок в Башкортостане. Башкирский орнитологический вестник. Уфа, РИО БашГУ. Вып. 3. 23-24.
Валуев В.А., Едрёнкин В.А. К антропогенным воздействиям на авифауну. Материалы III Международной научно-практической конференции. 12-13 мая 2005 г. Астрахань, издательский дом «Астраханский университет». 80-82.
Захаров В.Д. Птицы Южного Урала. (видовой состав, распространение, численность). Екатеринбург; Миасс: ИГЗ УрО РАН, 2006. 228 с.
Ильичёв В.Д., Фомин В.Е. (1988): Орнитофауна и изменение среды (на примере Южно-Уральского региона). М: Наука. 1-246.
Кириков С.В. Птицы и млекопитающие в условиях ландшафтов южной оконечности Урала. М., 1952, 1-412.
Коблик Е.А., Редькин Я.А., Архипов В.Ю. Список птиц Российской Федерации.М.: Товарищество научных изданий КМК. 2006. 256.
Кузякин А.П. Зоогеография СССР // Учён. зап. Моск. обл. пед. ин-та им. Н.К. Крупской. М., 1962. т. 109. С. 3-182.
Полежанкина П.Г. (2006). К редким видам птиц Башкирии. Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. Екатеринбург, изд-во Уральского университета. 168-169.
Равкин Ю.С. (1967): К методике учёта птиц в лесных ландшафтах. – Природа очагов клещевого энцефалита на Алтае. Новосибирск: Наука. 66-75.
Сушкин П.П. (1897): Птицы Уфимской губернии. Москва: изд-во Товарищество И.Н. Кушнеревъ и К°. 1-325.
Торгашов О.А. (2004). К фауне птиц национального парка «Башкирия». Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. Екатеринбург, изд-во Уральского университета. 178-182.
Эверсманн Э.А. Естественная история Оренбургского края. Оренбург: Изд-во ОГПУ, 2001. 344 с.
Fox G., Gilman A., Peakall D., Anderka F. (1978): Behavioral abnormalities of nesting lake Ontario Herring gulls. – J. Wildlife Manag.,vol. 42, N 3. 477-483.
С работами Валуева В.А. можно ознакомиться на сайте башкирского республиканского орнитологического общества http://www.broo.bashkiria.ru
Июн
19
Ответы на комментарии Полежанкиной П.Г., опубликованные на сайте института ЭкоЭксперт http://ecobioexpert.ru/.
Валуев В.А.
E-mail: ValuyevVA@mail.ru
// Башкирский орнитологический вестник. 2016. № 17. С. 47-50.
На нашем сайте появились комментарии Полежанкиной Полины Геннадьевны (в дальнейшем П.), цель которых, как можно понять из её высказываний, – опорочить некоторых членов Башкирского республиканского орнитологического общества (БРОО). Т.к. меня выбрали руководителем БРОО, то считаю своим долгом вступиться за честь людей, которые доверили мне руководство обществом.
Ниже приведены комментарии Полежанкиной П.Г. (выделено курсивом) на сайте института экологической экспертизы и биоинформационных технологий 13 января 2016 г. и наши ответы на них.
1 комментарий П.
«Эх, кто бы мне статьи из pdf в doc перевёл. Так чудненько копировать с других сайтов размещённую информацию. А раздел ваш, кстати, называется «Публикации (БРОО)», только вот Торгашов никогда отношения к БРОО не имел (за исключением публикации одной своей статьи в Баш. орнит. вестнике) и членом БРОО не являлся».
Ответ на комментарий № 1.
А) То, что П. сожалеет о нашем способе размещения своих сборников в pdf, уже говорит о том, что она занимается плагиатом – скачивает наш материал и размещает его на своём сайте http://xn—-8sbc0adaan4bqp3c3a2b.xn--p1ai/site/Stat-i-o-pticah/Nauchnye-stat-i/Po-avtoram. Мы уже указывали ей на допустимость таких действий, но она игнорирует это. Видимо, дожидается, когда мы подадим на неё в суд за плагиат.
Б) То, что Торгашов не является членом БРОО, – удар пальцем в небо; т.к. БРОО публикует данные о птицах республики независимо от принадлежности автора к какой-либо партии или обществу. Причём на сайте БРОО выложены и статьи самой П, хотя её исключили из членов БРОО ещё в 2014 г. Тогда, по её же мнению, следует и её статьи оттуда убрать? Пусть изволит желание – уберём.
2 комментарий П.
Приводя выдержку из статьи А.А. Романова (2015): «Близкое расположение территории урочища к крупному промышленному центру – Аша» П. рассуждает: «И как это понимать? Что все птицы из урочища Старошалашово перекочевали в Ашу, или что они все подохли или подальше улетели от Ашинского металлургического завода?»
Ответ на комментарий № 2.
А) В своей статье А.А. Романов указал на географическое расположение урочища Старошалашово – т.е. привязал его местонахождение к городу Аша, который легче найти на карте, чем само урочище. Что в этом криминального? Чем руководствовалась П. когда вопрошала “что, все птицы урочища перекочевали в Ашу”?
Б) Из чего П. взяла, что все птицы «подохли или улетели от Аши»? Видимо П. была переутомлена, когда это писала; т.к. в первом своём предложении она предполагает, что все птицы перекочевали в Ашу, а во втором – что они оттуда улетели.
3 комментарий П.
Анализируя статью А.Н. Кутлиахметова (2015) П. пишет: «Что-то маловато ссылок на Валуева. Глядишь, так резко индекс цитирования у последнего понизится. Из 25 статей, в которых имеется информация о совах (Валуев, 2002-2004, 2010, 2014-2014б; Валуев В., Валуев К., 2001, 2009, 2010, 2011; Валуев В.,Валуев Д., 2004; Романова, 2014).» Похоже что, например, Лоскутова, Алексеев, Бакалова вообще зря проводили свои многолетние исследования в заповедниках «Башкирский», «Южно-Уральский» и «Шульган-Таш». Всё = в итоге их даже не упомянули, как бы и не было их научных статей.»
Ответ на комментарий № 3.
А) А.Н. Кутлиахметов сделал обзор литературы, на основе данных исследователя, объездившего всю республику, чтобы на этой основе указать на редкость вида. По-моему это правильный подход, т.к. показывает картину существования вида в целом по республике. Если бы я, например, стал писать о чёрной нектарнице, которую видел в ЮАР и которая вообще не значится в списках птиц Ю. Африки, то тоже взял бы за основу труды того исследователя, который посвятил десятки лет своей жизни работе в этом регионе и не стал бы искать среди многочисленных конференций статьи тех людей, которые работали на этой территории, но нектарницами никогда не занимались.
Б) Лоскутова Н.М. (работала в РБ в 1980-х гг.), Лоскутова И.А. (в 1990-х работали в Башкирском заповеднике; Алексеев В.Н. изучал боровую дичь в Южно-Уральском; Бакалова М.В. – в Шульган-Таш. Эти заповедники составляют лишь часть горной области, о которой писал Кутлиахметов А.Н.; и в его задачи не входило подробное упоминание всех встреч с ястребиной совой на территории Башкирии. Что же касается упомянутых сотрудников заповедников, то они могут лишь подтвердить факт её присутствия в горах, в чём Кутлиахметов А.Н. и не сомневается – о чём так и написал; “этот вид гнездится лишь в горах”.
В) Если П. так сердечно воспринимает неупоминание какого-либо автора в работах других, то почему она не поставит это в упрёк В.К. Рябицеву, написавшего справочник о птицах Предуралья, Урала и Западной Сибири? Он не только не сделал ссылок на упомянутых П. авторов, но и на её же саму, Валуева, Фоминых и др. авторов написавших свои работы по птицам Башкирии? Почему П. не ставит вопроса о правомочности создания Красной книги РБ (2014) перед МПР РБ и правительством РБ в связи с тем, что очерки в ней писали люди не работавшие в РБ? Почему она направила свой яд на людей только, что вступивших на путь исследований в области орнитологии? Чтобы отбить у них желание заниматься этой наукой? Зачем?
Г) В своей статье “О сроках прилёта птиц …” (заметьте, за 2015 г.!) http://xn—-8sbc0adaan4bqp3c3a2b.xn--p1ai/site/Stat-i-o-pticah/Nauchnye-stat-i/Po-avtoram/Polezhankina-P.G/2015/O-srokah-priliota-nekotoryh-vidov-ptic-v-Respubliku-Bashkortostan П., кстати, сама не упоминает ни о Лоскутовых (Н.М., И.А., А.В.), ни об Алексееве В.Н., ни о Бакаловой М.В.. А ведь эти люди по своим должностям должны заниматься фенологией – прилётом и отлётом. Вот уж где уместны ссылки на работы этих авторов! Вот где следует П. повторить слова её же самой: “Похоже, например, Лоскутова, Алексеев, Бакалова вообще зря проводили свои многолетние исследования в заповедниках… Всё равно, в итоге их даже не упомянули, как и не было их научных статей“.
Литература:
Кутлиахметов А.Н. К распространению ястребиной совы Surnia ulula в Башкирии // Башкирский орнитологический вестник: сборник статей. Вып. 13. – Уфа: РИЦ БашГУ, 2015. С. 29-32.
Романов А.А. Авиафауна урочища «Старошалашово» (Ашинский район Челябинской области) // Башкирский орнитологический вестник: сборник статей. Вып. 13. – Уфа: РИЦ БашГУ, 2015. – С.32-36.
Апр
19
Клысова Д.Р.
Апр
19
Гибридизация Haliaeetus albicilla и H. leucoryphus?
УДК 598.279.23
Валуев В.А.
E-mail: ValuyevVA@mail.ru
// Редкие и исчезающие виды животных и растений Республики Башкортостан. 2016. № 11. C. 3-6.

