Хабибуллин В.Ф.
Амурский зоологический журнал, 2013. №4. С.381-390.
Ключевые слова: трофические группы животных, дедуктивный подход, трофический спектр, объект питания
Key words: animal trophic groups, deductive approach, trophic range, food object
Резюме. Трофические группы животных выделяются на основании специфики потребляемого объекта питания. При такой постановке вопроса задача выделения системы трофических групп животных сводится к задаче аде-кватного выделения в объеме исходного единого понятия «пищевого объект» системы подчиненных, меньших по объему понятий с применением операций логического и мереологического деления. При разбиении исход-ного понятия «пищевой объект» мы опираемся на различные признаки объекта питания: таксономическое по-ложение, особенности экологии или потребляемые части (соответственно таксономический, экологический и мереологический подходы). Таким образом, при дедуктивном подходе к созданию экологической классифика-ции животных (по характеру питания) мы всю систему строим «сверху» – дедуктивно выводя трофические группы из единого общего основания – понятия «пищевой объект» (объект питания).
Summary. In this paper we applied the deductive approach to distinguish the animal trophic groups and used the single concept “food object” as a basis. The “character of animal nutrition” is a qualitative composition and nature of con-sumed food objects. The multi-level hierarchical division of trophic groups can result in construction of ecological clas-sification of animals based on the “feeding character”. We used taxonomical, ecological and mereological approaches, in which we used as the base for division correspondingly the taxonomic status, ecological features and consumed parts of food objects. The “trophic range” was estimated on each level of division as the degree of food specialization by using the scale “stenophagy – euriphagy”. As a whole, the deductive approach is strongly logical and has such advantages as unified terminology and prognostic capability.
ВВЕДЕНИЕ
Трофические отношения считаются основополагающими как в отдельных биоценозах, так и в биосфере в целом [Шилов, 2003). Значение пищевых связей в биосфере настолько велико, что биотический кругооборот иногда рассматривают как трофологический процесс, а саму биосферу – как трофосферу, где пищевые связи образуют замкнутый круг [Уголев, 1980].
В экологии животных питание считается базовой функцией организма и рассматривается как основная фор-ма его связи со средой [Наумов, 1963]. На основе изучения питания выделяются трофические группы живот-ных, жизненные формы, создаются экологические классификации, изучаются пути приспособления организмов к среде [Чернова, Былова, 2007]. Во многом на основании именно изучения потребляемых пищевых объектов делаются выводы о роле животных в природе и значении для человека.
Необходимая сторона процесса познания – это огрубление конкретной действительности, разделение её на отдельные «сечения» и закрепление информации о них [Еганов, 1971]. Гносеологически проблема выделения объекта изучения сводится к процедурам отождествления и различения [Горский, 1984]. Выработка критериев различения связана с боль¬шими трудностями – ибо в силу диалектического характера дейст¬вительности между объектами не существует строгих разграничительных линий; объекты как материальные явления подвижны, изменчивы; объекты из числа абстрактных объектов науки зависят от характера соответствующей области знания, принимаемых допущений и т.п.
Выделение объекта из многих других должно быть четким и однозначным – без этого невозможно их иссле-дование. Субъекту познания представляется весьма богатый выбор возможностей вычленения материальных объектов по разнообразным наборам признаков. Весь вопрос в том, какую из этих возможностей и из каких соображений он реализует» [Еганов, 1971]. Сложности, связанные с отождествлением предметов с самими со-бой и друг с другом, проведение конструктивизирующих, «жестких» границ между множествами предметов посредством их определений заставляет прибегать к идеализациям, допущениям и оговоркам [Горский, 1984]. Получается, что любое изучение объекта возможно только в ситуации идеальных границ, которых реально не существует.
В ходе изучения трофики животных накоплено значительное количество эмпирического материала, для систематизации которого применяются методы группировки и классификации. В большинстве случаев осуществляется именно группировка имеющихся данных по «характеру питания» (который понимается очень широко – от способа добывания пищи до характеристик потребляемых объектов). Такие объединения не могут считаться классификациями (ибо не удовлетворяют требованиям, предъявляемым к подобного рода построениям), но прагматически полезны, т.к. способствуют решению поставленных задач – характеризуют изучаемые аспекты питания животных.
Основная цель (и главная функция) классификации – обеспечить однозначное и легко определимое место для каждого из классифицируемых объектов. Она должна одинаково хорошо выполняться при двух основных видах использования классификации: размещении нового объекта в классифицируемом массиве и нахождении конкретного объекта в этом массиве [Кузнецов, Титов, 1998]. Из существующих методов классификаций широко применяются фасетный (каждая выделенная группа характеризуется собственным набором признаков) и иерархический (между выделенными группами устанавливаются отношение подчинения).
Строить классификацию можно двумя способами: индуктивным и дедуктивным. В большинстве экологических работ применяется индуктивный способ. Исходным материалом для создания классификации служит массив неорганизованных данных (= вариантов классифицируемых объектов) – членов нижнего уровня будущей классификации, в т.ч. с примесью посторонних. Чтобы выявить и отсечь лишнее, нужно поставить вопрос: «Выделен ли данный вариант классифицируемых объектов на основании признака «объект классификации»? При этом важно определиться с обобщенным названием объекта классификации (= вершинной ячейки), т.е. что именно мы классифицируем, а также четко представлять основание классификации – особенности или варианты признака, присущего данным предметам, по которому производится выделение классификационных вариантов. Так, не следует смешивать трофические группы, формы межвидовых трофический связей или трофические уровни (например, «гематофаг», «паразит» или консумент), как выделенные по разным основаниям. После составления (выделения) перечня трофических групп путем последовательного объединения создается многоуровневая классификация животных по характеру питания.
Достоинством индуктивного подхода является то, что все выделяемые трофические группы содержательны, т.е. имеют фактическое наполнение. Однако полученная индуктивным путем классификация целиком определяется экологией исследованных объектов (в т.ч. степени их изученности), имеет свою специфическую «локальную» терминологию, не может без серьезных доработок быть применена к другим группам животных, не обладает прогностической способностью. С точки зрения логики при индуктивном варианте построения классификации приходится использовать две вариантообразующих операции: деление и обобщение (с подбором основания и попарного сравнения всех вариантов на каждом уровне), поэтому алгоритм работы достаточно сложен и трудно поддается формализации [Кузнецов, Титов, 1998].
Указанных недостатков лишен дедуктивный подход, применение которого для выделения трофических групп животных мы рассмотрим в данной работе. Классификации трофических групп животных будет проводиться с опорой не на наблюдаемые феномены, но на основе вариации признака, лежащего в основании классификации – объекта питания [Хабибуллин, 2012].
МЕТОДОЛОГИЯ И ЛОГИКА ДЕДУКТИВНОГО ПОДХОДА К ВЫДЕЛЕНИЮ ТРОФИЧЕСКИХ ГРУПП И ИХ КЛАССИФИКАЦИИ
Дедуктивный способ в качестве отправной точки имеет единственный объект – основание (объект) классификации; в данной работе таковым является понятие «пищевой объект (объект питания)», под которым пони-мается органическое вещество любого происхождения, состава и структуры; отдельные организмы любой систематической принадлежности, строения и образа жизни, любые части особей, их прижизненные выделения и посмертные останки. Субъект питания – отдельное животное-потребитель пищевого объекта или трофическая группа (совокупность животных, выделенная на основе потребления объекта питания).
Методологические положения, принятые нами в качестве исходных, таковы:
1. Животные – гетеротрофные организмы, питающиеся органическим веществом – совокупностью пищевых объектов.
2. Все пищевые объекты обладают свойствами фагичности (доступность для животных-потребителей в качестве источника пищи) и трофичности (способность быть ассимилированным организмом животного-потребителя).
3. Качественный состав и природа потребляемых пищевых объектов характеризуют характер питания животных (=пищевой режим, рацион, диета; пищевая специализация первого порядка [Бей-Биенко, 1980].
4. Трофические группы животных выделяются на основании потребляемых объектов питания, без учета способов их добывания, поглощения, переваривания, типов межвидовых взаимодействий и пр.
5. В качестве основания для выделения отдельных групп объектов питания правомерно использовать любые их признаки: таксономическое положение, экологические особенности, анатомо-морфологическое строение.
6. Выделять трофические группы можно для любой совокупности животных: как для таксоценов различного уровня, так и для любых экологических групп.
7. Многоуровневое иерархическое выделение трофических групп позволяет создать экологическую классификацию животных по характеру питания.
8. Трофический спектр (=пищевая специализация второго порядка [Бей-Биенко, 1980]) – многообразие потребляемых пищевых объектов, отображает степень пищевой специализации по шкале «стенофагия – эврифагия»; оценивается на одноуровневой ступени классификации.
В основе дедуктивного способа формирования классификации лежит единственная логическая операция – «деление понятия». Деление есть разбиение множества объектов, составляющих объем делимого понятия, на непересекающиеся подмножества (члены деления) по выбранному единственному на каждом шаге деления основанию деления понятия. Подчеркнем, что трофические группы, выделенные по основанию «потребляемый объект питания», отличаются от сходных образований, выделенных по другим основаниям: функциональных трофических групп – соскребатели, собиратели и пр. [Gaines et al., 1989], функционально-трофических групп – зооризомицетофаги, филлонекрофитофаги и пр. [Второв, Дроздов, 2001], функциональных биоценотических групп – филлофаги, зоопаразитоиды и пр. [Панфилов, 1966]. Операция деления включает в себя: 1) делимое понятие; 2) основание деления; 3) результаты деления (члены деления) [Ивлев, 2002]. Помимо операции собственно логического (таксономического) деления выделяют так называемое мереологическое (аналитическое) деление как процедуру мысленного расчленения предмета на составные части [Ивин, 2000]. Для целей нашей работы мы будем использовать обе операции деления.
Классификация (иерархическая) понимается как многоступенчатое деление логического объема понятия на систему соподчиненных классов объектов. Под уровнем деления (уровнем иерархической классификации) понимается этап (шаг, ступень) в многоступенчатой операции деления; под глубиной деления – количество иерархических уровней деления. Объем исходного понятия (вся мыслимая совокупность пищевых объектов) мы далее в ходе последовательных делений разбиваем части (по которым образуем трофические группы животных), иерархическая многоуровневая система которых образует экологическую классификацию животных по характеру питания. Полное число ячеек последнего (нижнего) уровня составляет объем классификации.
При выделении трофических групп важно соблюдать общие правила логического деления:
Правило единственного основания: на каждой ступени деление должно вестись только по одному основанию. Ошибка: деление по разным основаниям, когда члены деления выделены по различным признакам.
Правило соразмерности: деление должно быть соразмерным, или исчерпывающим, т.е. сумма объемов членов деления должна равняться объему делимого понятия. Ошибка: неполнота деления, когда выделенные члены деления в сумме не покрывают всего объема делимого понятия.
Правило разграниченности: члены деления должны взаимно исключать друг друга. Ошибка: деление неисключающее, когда члены деления пересекаются.
Правило непрерывности: деление должно быть непрерывным. Ошибка: скачок в делении, когда основание деления одно, но некоторые из его членов выделены с учётом некоторого добавочного признака.
В ряде случаев трудно быть уверенными в том, что мы выделили все возможные варианты, поэтому на каждом этапе деления можно предусмотреть наличие ячейки «прочие». В некоторых случаях выделенные логическим путем ячейки окажутся фактически пустыми (т.е. потенциально пищевой объект есть, а его потребителя – нет), но могут получить содержательное наполнение в будущем. Элементы индуктивного подхода присутствуют в том смысле, что исследователь до начала создания классификации имеет некоторое преставление об объектах питания и возможных трофических группах. Может возникнуть вопрос об обоснованности дедуктивно выделенных трофических групп. Например, есть ли животные, которые питаются только плаунами? Или только запасающей тканью растений? Или только щупальцами кишечнополостных? Мы считаем, что даже один-единственный трофически специализированный потребитель «заслуживает» выделения отдельной ячейки в трофической схеме и отдельного термина. Кажущееся неочевидным или даже излишним выделение от-дельных трофических групп при более внимательном рассмотрении оказывается вполне обоснованным и эмпирически: например, выделение потребителей линочных шкурок личинок и экзувиев куколок членистоногих, объем которых в широколиственных лесах или луговых степях огромен [Чернов, Руденская, 1975].
Фактуальное заполнение дедуктивной схемы эмпирическими данными производится как в процессе ее создания, так и позднее, при изучении питания конкретной группы животных. Такой подход позволяет устранить недостатки индуктивного подхода: обеспечивает единообразие терминологии, последовательно охватывает все множество пищевых объектов, обосновывает использование логических оснований для их деления, упорядочивает процедуру выделения трофических групп, обладает предсказательной способностью.
В ходе выполнения работы мы применяли прагматический подход [по Б.М.Миркину и др., 2001], с изменениями), включающий следующие основные положения:
необходимость – нужно выделять трофические группы для животных с самыми разнообразными пищевым режимом и трофической специализацией;
доступность – основания для выделения трофических групп должны быть доступны для выявления;
информативность – основания, по которым выделяются трофические группы, должны отражать существенные особенности пищевых объектов с точки зрения их фагичности и трофичности;
единообразие – использование единой терминологии, унификация принципов выбора оснований для выделения трофических групп животных;
оптимальность – уровень выделения и объем трофических групп должен быть удобным для практического использования.
ТРОФИЧЕСКИЕ ГРУППЫ
Широкая трактовка трофики [Уголев, 1980] рассматривает процесс питания как потребление не только органических, но и неорганических веществ (предложены соответствующие термины: биотрофия и абиотрофия). Заметим, что термин «абиотрофия» фактически означает питание неорганическими веществами независимо от их «биологичности»; в термине же «биотрофия» приставка «био-» подразумевает биологическое происхождение потребляемых органических веществ, упуская из виду органику небиологического, в том числе антропиче-ского происхождения. Исходя из этих рассуждений мы считаем, что корректнее использовать термины «органикотрофия» и «инорганикотрофия» (=абиотрофия) для обозначения потребления соответственно органических и неорганических веществ; и в ходе дальнейших рассуждений мы будем считать, что все животные – «органикотрофы» = «гетеротрофы». Термины же «витальная экзотрофия» и «поствитальная экзотрофия» (в смысле [Уголев, 1980]) предлагаем считать синонимами терминов «биофагия» (=биотрофия) как питание живой органикой и «некрофагия» как питание неживой органикой. Грань между биофагией и некрофагией тонка. Организмы, поедающие живые «части» тела объекта питания – биофаги, организмы, умерщвляющие «часть» тела живого хозяина и поедающие уже мертвую органику, находящуюся в живом теле объекта питания – некрофаги. В строгом смысле некрофагами следует считать и хищников; градации же объекта питания по степени «свеже-умерщвленности» варьируют вплоть до сильно разложившейся органики.
При образовании терминов для именования трофических групп используется греческий или латинский (ре-же – русский) корень от названия объекта питания, с прибавлением окончания «-троф», «-фаг» или «-ядный». Их использование неравномерно в различных разделах биологии: окончание «-фаг» традиционно применяется, например, в энтомологии и герпетологии, «-троф» – в физиологии и гидробиологии, «-ядный» – в орнитологии, териологии. Из этих окончаний первое представляется нам наиболее предпочтительным как семантически ней-тральное и универсальное, тогда как второе имеет дополнительную смысловую нагрузку (подразумевает по-глощение относительно крупных частей) и лучше отражает сущность питания животных, их деятельное начало – в противоположность пассивной «трофности». Русские же названия часто неблагозвучны, как-то [Пономарева, 1975]: цветкоеды, почкоеды, сокососы. В целом мы придерживались сложившейся терминологии, предложив лишь несколько операциональных терминов. Значение многозначных терминов необходимо оговаривать. Например, термин «филлофагия» иногда расширенно трактуется как потребление фотосинтезирующих частей растений независимо от того, являются ли такие части растений листьями или нет [Пономаренко, 2006], что по сути равнозначно «хлорофагии» как потреблению асиммиляционной ткани [Рожков, 1981]; встречается и расширенное трактовка ксилофагии как потребление луба, коры и древесины [Линдеман, 1980]. На наш взгляд, оба термина следует трактовать узко: филлофагию – как потребление листьев (органа, а не ткани), ксилофагию – как потребление древесины (комплекса тканей, а не органа).
Логические аспекты дедуктивного выделения трофических групп животных суммированы в таблице, где четко указаны: делимое понятие, основание деления, результат деления. В четвертом столбце приведены в ка-честве примеров названия некоторых трофических групп.
Таблица
Логические аспекты выделения трофических групп животных
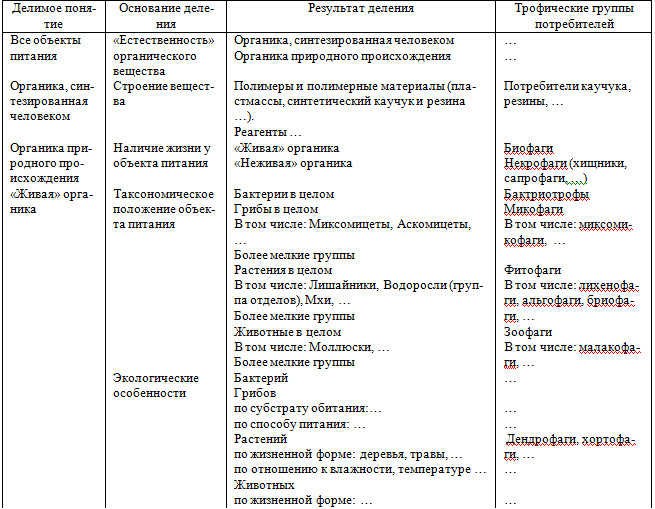


Основание классификации должно быть детерминировано признаком, существенным для решения задачи с помощью данной классификации (гносеологические требования существенности основания). Без указания ос-нования деления понятия (основания классификации) нет и классификации; – в лучшем случае есть простое перечисление некоторых подмножеств определенного множества объектов [Петров, 1990]. В зависимости от целей работы исследователь вправе выбрать любое основание деления, которое он считает необходимым; в нашем случае нужно использовать существенные в трофическом отношении признаки, от более общих до част-ных.
Схема выделения трофических уровней приведена на рисунке 1.
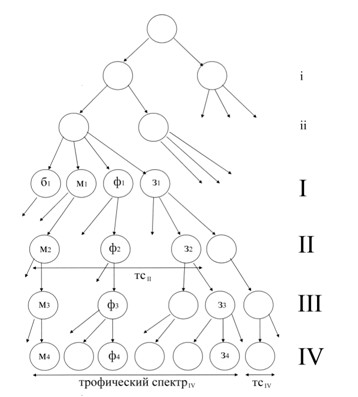 Рис. 1. Схема выделения трофических групп и определения ширины трофического спектра.
Рис. 1. Схема выделения трофических групп и определения ширины трофического спектра.
Исходное делимое понятие (верхний круг) – вся совокупность объектов питания. i-ii – предварительные уровни деления пищевых объектов и выделения трофических групп животных: i – объекты питания природного или антропогенного происхождения; ii – объекты питания «живые» или «неживые». I-IV – основные уровни деле-ния; в качестве примера приведено деление по таксономическому основанию: I – объекты питания на уровне царств и их потребители (б1 – бактериотрофы, м1 – микофаги, ф1 – фитофаги, з1 – зоофаги); II – объекты пита-ния на уровне типа/отдела и их потребители; III – объекты питания на уровне класса и их потребители; IV объекты питания на уровне отряда/порядка и их потребители. Примеры фактического заполнения ячеек м2-м4, ф2-ф4 и з2-з4 приведены в тексте. Показана ширина трофического спектра на втором уровне (тсII) и на четвертом уровне (трофический спектрIV и тсIV).
На предварительных уровнях деления i-ii использованы единые общебиологические основания для выделе-ния объектов питания. На первом предварительном уровне деления (i) органическое вещество как объект пита-ния можно подразделить на «естественное» (природного происхождения) и «искусственное» (синтезированное человеком).
На втором предварительном уровне деления (ii) искусственную органику как объект питания можно подраз-делять на группы, руководствуясь классификатором продуктов органического синтеза [Москвичев, Фельд-блюм, 2007]. В качестве примера потребителя такой синтетики можно привести хлебного точильщика Stego-bium paniceum который «ест все, кроме железа» [Бей-Биенко, 1980], в т.ч. сухие лекарственные средства. Син-тезированные химической промышленностью вещества разнообразны (миллионы наименований) и обильны. С экологической точки зрения появление новых органических веществ означает появление исторически нетипич-ных пищевых ресурсов, освоение (потребление) которых животными возможно и непосредственно, но чаще связано с приобретением и культивированием соответствующих эндосимбионтов из числа бактерий, грибов или простейших. В таких условиях отбор может идти по пути создания устойчивой системы «животное – эндо-симбионт», способной утилизировать появившийся пищевой ресурс, при этом в метаболитическом и особенно трофическом отношении животные как ассимилирующий организм вместе с их эндосимбионтами рассматри-ваются как единое целое, как форма образования надорганизменных генетических систем [Тихонович, Прово-ров, 2009]. Присутствие «случайных» загрязняющих бактерий, способных расщеплять нехарактерные для дан-ного потребителя пищевые субстраты может рассматриваться как приспособление к новым трофическим ни-шам [Уголев, 1991].
Объекты питания природного происхождения (на том же втором предварительном уровне деления ii) можно подразделить на «живые» и «неживые». Граница между живым и неживым не всегда четко выражена, и задача их разграничения не так проста, как может показаться на первый взгляд; это касается в первую очередь частей отдельных организмов, как внутренних (например, древесина), так и внешних (например, перья, волосы, ли-стья), в том числе и различных прижизненных выделений. Мы предлагаем опираться на следующий формаль-ный критерий: если объект питания – часть живого существа, или находится непосредственно на/в нем, то он считается «живым» (например, сосуды древесины, листья в состоянии «листопадности», частицы эпителия, пот и пр.); если же пищевой объект находится вне живого существа, то он считается «неживым». Для группы по-требителей прижизненных выделений мы предлагаем использовать термин «ретриментофагия» (от латинского retrimentum – отходы, выделения, экскременты). В широком смысле ретриментофагия охватывает потребление всех организменных выделений – от пота, слез и нектара до мочи и экскрементов и включает в себя среди про-чих таких потребителей, как копрофаги, кератофаги (часть), нектарофаги. Здесь же логично рассматривать термины «лакрифагия» – потребление слезных выделений, подсасывание пота и других железистых выделений [Балашов, 1982], и «мукофагия» – потребление слизи, гноя.
На всех основных уровнях деления выбор основания для разделения пищевых объектов и выделения трофи-ческих групп жестко не фиксирован и в качестве основания логической операции деления могут выступать: таксономическое положение объекта питания, особенности экологии объекта питания, потребляемые части объекта питания. Соответственно используется таксономический, экологический и мереологический подходы.
При таксономическом подходе встает вопрос о принятии того или иного из существующих вариантов систематики пищевых объектов. Сложности возникают уже на уровне царств: так, обсуждается самостоятельность царств простейших Protista, так называемого «среднего» царства Protoctista, царства Chromista (включающее оомицетов) и т.п. [Мирабдуллаев, 1989; Мухин, Третьякова, 2007 и др.]. Мы предлагаем придерживаться консервативной точки зрения, и для целей нашей работы в качестве объектов питания животных прагматично рассматривать четыре царства: Бактерии (Bacteria + Archaea), Грибы (в том числе грибообразные организмы), Растения (включая водоросли и лишайники), Животные (одно- и многоклеточные). В соответствии с данным походом выделяются и трофические группы животных: бактериотрофы (термин «бактериофаг» преоккупирован вирусами – пожирателями бактерий), мико-, фито-, и зоофаги. Отметим широкую распространенность бактериотрофности среди водных и почвенных животных (в первую очередь среди одноклеточных, но не только) – так, среди почвенных нематод доминируют именно бактериотрофы, потребляющие до 25% урожая бактерий [Покаржевский, 1985]. В целом таксономический подход удобнее для применения в том смысле, что имеются «готовые», более-менее общепризнанные, выделенные по таксономическому принципу группы пищевых объектов.
При экологическом подходе ввиду огромного разнообразия экологии объектов питания мы предлагаем отбирать лишь признаки, важные для целей выделения трофических групп. Это могут быть самые разные экологические черты объектов питания: жизненные формы (в том числе выделенные по отношению к отдельным абиотическим факторам – свету, температуре, субстрату), стадии онтогенеза, размеры, образ жизни, в том числе его отдельные пространственные и временные аспекты, поведенческие черты и многое другое. Примеры объектов питания, выделенных при экологическом подходе: дендробионты, яйца птиц, стволовые личинки насекомых, мезобентос, грибки хвойных пород, животные с сумеречной активностью, колониальные обрастатели, организмы с положительным фототаксисом. Содержательное наполнение эти группы получают при соотнесении их с животными – потребителями. Этот подход пока слабо разработан, но представляется перспективным как отражающий экологическую сущность выделения трофических групп.
При мереологическом подходе для выделения частей пищевых объектов можно отталкиваться от их морфологии или гистологии. В первом случае выделяют части тела, органы и их системы; во втором – отдельные ти-пы клеток, ткани или их комплексы. Мы предлагаем дополнительно к известному термину «голофагия» – потребление пищевого объекта целиком, использовать термин «партифагия» – потребление отдельных частей пищевого объекта. Несколько особняком должны рассматриваться варианты питания содержимым кишечника (например, цестоды) или органическим ассимилятом, текущим по флоэме высших растений (например, тли). В обоих случаях потребляемая «живая» органика формально может считаться частью пищевого объекта. Практическим преимуществом мереологического подхода является возможность использования достижений гистологии и морфологии отдельных групп организмов – пищевых объектов [см., например, О.А.Коровкин, 2007].
На каждом уровне можно применять любой из трех упомянутых подходов; возможно и выделение трофических групп животных с использованием только одного подхода.
Последовательно применяя таксономический подход, можно дойти до выделения отдельного вида – пищевого ресурса для узкоспециализированной группы животных. При этом в качестве основания для выделения трофических групп не учитываются особенности экологии или мереологии пищевых объектов. В идеале (но не обязательно) могут быть даже соблюдены следующие условия: 1) экологическая однородность объекта питания – т.е. исключение вычленения отдельных экологических групп пищевых объектов; 2) потребление объекта питания целиком (голофагия) – т.е. исключение мереологического вычленения отдельных поедаемых частей. В результате мы получаем таксономически монофагичную группу. Монофагичность – свойство трофической группы, потребление однородных (таксономически, экологически, мереологически) объектов питания.
Возможен и абсолютный экологический подход: выделение группы животных, потребляющих один и толь-ко один экологически однородный пищевой объект (стадию онтогенеза, размерную группу, жизненную форму и т.п.) – например, потребители птичьих яиц. Идеальные условия аналогичны: 1) независимость от таксономической принадлежности объекта питания; 2) голофагия. В результате мы получаем экологически монофагич-ную группу.
Возможен и абсолютный мереологический подход: выделение группы животных, потребляющих одну и только одну часть пищевого объекта (один тип клеток, одну ткань, один орган, одну часть тела) – примеры уз-кой гостальной специфичности нам предоставляет паразитология. Идеальные условия: 1) независимость от таксономического положения объекта питания; 2) экологическая однородность объекта питания. В результате мы получаем мереологически монофагичную группу.
Таксономические и отчасти мереологические различия потребляемых пищевых объектов означают различия в их трофичности для животных-потребителей: со стороны объекта это особенности биохимии, морфологии, структуры; со стороны субъекта – строение желудочно-кишечного тракта в целом и ротового аппарата в частности, наличие или отсутствие ферментов для расщепления соответствующей пищи, эффективность усвоения и т.п. Экологические различия потребляемых пищевых объектов означают различия в их фагичности: доступность пищевых объектов для животных-потребителей определяется экологией обоих участников: их образом жизни, распространением в пространстве и времени, всем спектром взаимодействий: защитных и атакующих приспособлений, способами добывания пищи (в широком смысле: поиск, извлечение, захват, «выращивание» и пр.) и др.
Объяснить трофичность и фагичность тех или иных пищевых объектов, существующее положение с трофи-ческими группами и шириной трофических спектров можно посредством обращения к концепции [по Криво-луцкому, Покаржевскому, 1986, с изменениями] биологических барьеров, под которыми понимается любая преграда биологического характера, препятствующая потреблению пищевого объекта. В процессе добывания и усвоения пищевых объектов для животных-потребителей имеют значение следующие биологические барьеры: мембранные – как избирательная проницаемость мембран для нутриентов; физиологические – как избирательное потребление соединений из пищи и усвоение в желудочно-кишечном тракте, анатомо-морфологические – как специфическое обращение с объектом питания и, например, избирательное поедание отдельных его частей; биоценотические – как образ жизни, «адрес и профессия» объект-субъектной пары в ценозе и, следовательно, избирательное потребление пищевых объектов.
ТРОФИЧЕСКАЯ СПЕЦИАЛИЗАЦИЯ
Трофический спектр характеризуется многообразием пищевых объектов и через показатель степень (шири-ну) отображает пищевую специализацию потребителей.
Термины «моно-», «олиго-» и «полифагия» (абсолютная шкала) характеризуют трофическую «статику» – констатация дискретного состояния, динамичную же составляющую трофического спектра – континуальность – лучше отражает относительная шкала «стенофагия (= монофагия + олигофагия) – эврифагия» как отражение специализированности и неспециализированности в питании.
Трофический спектр можно оценивать для трофических групп, выделенных по разным основаниям, и тогда мы получим разное наполнение термина «монофагия»: при таксономическом подходе монофагией может считаться потребление пищевых объектов, относящихся исключительно к одному таксону (виду, роду, семейству, отряду и т.п.); при экологическом подходе – потребление только одной жизненной формы; лишь одной стадии онтогенеза и т.п.; при мереологическом подходе – потребление одной (и только одной) части тела, одного типа клеток/тканей/органов.
При установлении степени детальности (с точностью до какого уровня мы определяем монофагичность) важно учитывать «масштаб» [Levin, 1992; Schneider, 2001], который зависит от поставленных исследователем задач и «размера» анализируемой трофической группы. Вопрос об объеме понятия «монофагия», решается по-разному в разных группах животных. Наиболее строгие требования к монофагичности предъявляются к фитофагам: так, для растительноядных насекомых [Бей-Биенко, 1980]: монофагия – это потребление растений одно-го или нескольких видов в пределах одного семейства, олигофагия – потребление растений разных видов из одного или нескольких семейств, полифагия – потребление растений из многих семейств. Для зоофагов требования слабее и масштаб другой: например, считающая трофически узко специализированной обыкновенная медянка Coronella austriaca Laur., 1768 [Дробенков, 1995] питается различными видами ящериц (т.е. объектами в ранге отряда); среди птиц [Ильчев и др.,1982] монофагами считаются бакланы, питающиеся только рыбой – т.е. объектами в ранге надкласса.
Важно помнить, что на каждом уровне деления существует своя шкала «стено- – эврифагии», поэтому нужно предварительно оговаривать, на каком уровне (шаге деления) мы оцениваем широту трофического спектра и для какой именно группы потребителей.
Эврифагию можно понимать как совокупность монофагий более низкого уровня; с другой стороны, любая эврифагичная группа при продвижении «вверх» по классификации становится монофагичной. С этих позиций трофический спектр можно оценивать и аддитивно – как сумму трофических спектров более низкого уровня. Например, божьи коровки (жуки семейства Coccinellidae) на уровне семейства эврифагичны: среди них есть мико-, фито- и зоофаги. Такая «всеобщность» кокцинеллид как трофической группы – в терминах логики – объясняет слишком многое, и эту эврифагичность можно преодолеть, спускаясь на более низкие уровни выделения трофических групп, характеризуя уже эти «дочерние» трофические группы, а через них – аддитивно – исходную. Поэтому следующим шагом выступает дальнейшее деление: например, энтомофагов можно подразделить на афидофагов и кокцидофагов. Разнообразие насекомых как пищевых объектов в диете кокцинеллид некорректно характеризовать термином «миксоэнтомофагия» (как потребление насекомых из разных таксонов), т.к. он характеризует как раз ширину трофического спектра (=эврифагия), а не пищевой режим. На рисунке 1 можно проследить пошаговое деление пищевых объектов кокцинеллид: для микофагов на уровнях м2-м4: Отдел Настоящие грибы Eumycota – Класс Сумчатые грибы Ascomycetales – Порядок Мучнисторосяные грибы Erysiphales; для фитофагов на уровнях ф2-ф4: Отдел Покрытосеменные Angiospermae – Класс Двудольные Dicotyledones – Порядок Гвоздичные Caryophyllales; для зоофагов на уровнях з2-з4: Тип Членистоногие Arthropoda – Класс Паукообразные Arachnida – Отряд Акариформные клещи Acariformes.
Трофический спектр может быть узким (тсIV на рис. 1) или широким (тсII), сплошным – т.е. охватывающим смежные пищевые объекты (тсII на рис. 1) или дизъюнктивным, когда потребляются не смежные пищевые объ-екты (трофический спектр IV); в последнем случае иногда говорят об узко- или широкодизъюнктивном трофи-ческом спектре [Хрисанова, 2006].
АЛГОРИТМ ВЫДЕЛЕНИЯ ТРОФИЧЕСКИХ ГРУПП
Глубина деления (количество этапов деления) определяется достижением уровня с приемлемой стенофагичностью (в предельном случае – монофагичностью) (рис. 2): на каждом шаге деления мы проверяем вновь полу-ченную трофическую группу на монофагичность до тех пор, пока не будет достигнут такой объем понятия «пищевой объект», который был бы достаточным для обоснования выделения монофагичной группы живот-ных-потребителей. Заметим, что монофагичность может быть достигнута далеко не всегда: невозможно это, например, для эврифагичных низкоранговых таксонов. Весьма желательно, чтобы в разных ветвях глубина деления была одинакова. Группу, где монофагичность достигнута на более высоком уровне, можно довести до более низкого уровня, используя операцию «ограничение понятия». В рассматриваемом примере понятие «паукообразные» можно ограничить до понятий более низких уровней: «акариморфные клещи» и «паутинные клещи».
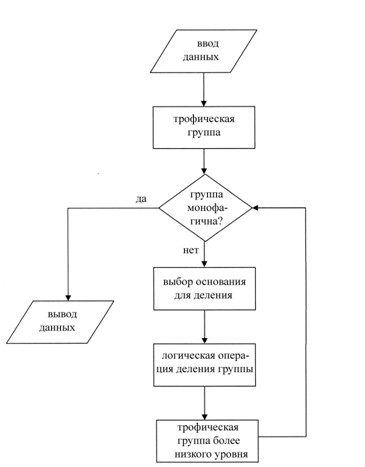 Рис. 2. Блок-схема алгоритма достижения монофагичности трофической группы.
Рис. 2. Блок-схема алгоритма достижения монофагичности трофической группы.
Добавить комментарий