В.Ф.Хабибуллин
Зоологический журнал – 2002. – Т. 83. – № 3. – С. 242-249.
Обсуждаются вопросы происхождения и формирования современной фауны пресмыкающихся Башкирии (10 видов). Дана краткая характеристика развития климата, ландшафтов и растительности. Приведены обобщенные палеонтологические данные по ископаемым позднекайнозойским пресмыкающимся из 7 местонахождений Башкирии (Южный Урал, Россия) и прилежащих территорий; определены 5 видов пресмыкающихся. Огромное влияние на развитие герпетофауны оказали плиоцен – плейстоценовые оледенения. Современная фауна пресмыкающихся Башкирии начала формироваться в конце плиоцена и окончательно сложилась к середине голоцена за счет расселения рептилий из рефугиумов. Уральские горы, как и антропогенные факторы, в историческом плане существенно не повлияли на процесс становления со-временной фауны пресмыкающихся Башкирии.
Современная фауна пресмыкающихся Башкирии насчитывает (Хабибуллин, 1999а) десять видов: один вид черепах, три вида ящериц и шесть – змей. Некоторые вопросы формирования герпетофауны Волжско-Камского края, в том числе и Предуралья Башкирии, обсуждались в работе Гаранина (1983); другой конкретной информации по этому вопросу мы не обнаружили.
Башкирия (Республика Башкортостан, РБ) расположена на Южном Урале и подразделяется на три зоны: Предуралье, горно-лесная зона и Зауралье. История формирования фауны пресмыкающихся Башкирии тесно связана с историей развития всей фауны Уральского региона и прилегающих территорий Евразии, а также с историей фауны Палеарктики в целом. Для разработки и создания общей картины мы не можем ограничиться только территорией Башкирии, в ряде случаев будем привлекать данные по смежным регионам. При изучении вопросов происхождения, развития и формирования современных фаун преимущественно используют два подхода: палеонтологический – на основе анализа палеонтологического материала и последующих палеозоологических экстраполяций, и ареалогический (зоогеографический), основанный на анализе современных ареалов ныне живущих видов. Используя данные палеонтологии, палеогеографии и зоогеографии, мы попытаемся предложить схему формирования современной фауны пресмыкающихся Башкирии.
КРАТКАЯ ХАРАКТЕРИСТИКА КЛИМАТА, ЛАНДШАФТОВ И РАСТИ-ТЕЛЬНОСТИ ЮЖНОГО УРАЛА В ПОЗДНЕМ КАЙНОЗОЕ
Современные ландшафты и природные зоны Башкирии очень разнообразны, что в том числе обусловлено и спецификой исторического развития климата, рельефа и ландшафтов Южного Урала. Природная среда обитания во многом определяет направление и характер развития фаун, поэтому рассмотрим основные этапы развития и формирования современных климата, ландшафтов и растительности на территории Южного Урала в позднем кайнозое.
Ранний кайнозой отличался равномерно теплым и стабильным изотермическим климатом; Южный Урал входит в так называемую квазитропическую зону. В результате среднекайнозойского похолодания в северном полушарии произошел распад квазитропического климата. В позднем олигоцене на пространствах зародившегося умеренного пояса (возможно, впервые и на Южном Урале) стал ложиться и некоторое время удерживаться снежный покров, способствовавший дальнейшему выхолаживанию и иссушению атмосферы; широкое распространение на большей части территории Палеарктики получают аридные условия (Синицын, 1980).
Основа современного рельефа Южного Урала формируется к концу миоцена (Яхимович, 1992), в это же время закладывается современная гидрографическая сеть с двусторонним стоком.
В плиоцене континентальность и сухость климата на юго-востоке Русской платформы значительно возросли. Горы Южного Урала в среднем плиоцене были покрыты хвойными лесами; покров травянистых и водно-болотных растений был очень богат. В отложениях нижнего плиоцена господствует уже почти безраздельно пыльца травянистых и кустарниковых растений (Немкова, 1981). Основы современного расположения растительных зон в Предуралье были заложены уже в раннем плиоцене.
Для четвертичного периода мы придерживались стратиграфической шкалы, применяемой для западного склона Южного Урала по изданию: Антропоген Южного Урала, 1965.
В нижнем эоплейстоцене (акчагыл) в связи с повышением уровня Каспийского бассейна два раза отмечалось появление моря в Предуралье Башкирии (Кайнозой Башкирского Предуралья, 1970). Во время раннеакчагыльской ингрессии моря при кратковременном по продолжительности, но максимальном по глубине погружении в Предуралье происходит проникновение бореальных вод с севера. Таким образом, в раннем эоплейстоцене в течение короткого промежутка времени Урал был отрезан от Русской равнины. В среднем акчагыле развивается максимальная ингрессия моря, воды проникают далеко в Башкирское Предуралье по системе древних долин: один залив с северо-запада по долине Палео-Белой; другой с юга – по долине Палео-Сакмары и ее притоков; соединению этих двух морских солодноводных заливов препятствовали возвышенности Общего Сырта (Антропоген …, 1965). Следовательно, в это время Южный Урал был отрезан от Русской равнины с северо- и юго-запада, к югу также располагалось море.
Южный Урал и его предгорья не покрывались ледником даже в эпоху максимального (днепровского) оледенения Русской платформы, лишь на высоких горных массивах и хребтах Южного Урала (а высоты гор были близкими к современным) в среднем и позднем плейстоцене присутствуют небольшие ледниковые отложения горно-долинного типа (Антропоген …, 1965), хотя существует мнение (Монин, Шишков, 1979), что для Урала именно окское, а не днепровское оледенение являлось максимальным: южная граница ледниковых морен достигала 55° с.ш., т.е. получается, что весь север Башкирии до широты Павловского водохранилища был покрыт ледниками.
В днепровское и московское время растительность на Южном Урале и прилегающих к нему равнинах была очень бедной, с множеством спор мхов, папоротников и обильными остатками грибов, воссоздающих картину перигляциальной степи (Антропоген …, 1965); большие пространства были заняты сфагновыми болотами и тундрами. Мощные снежники не только в горной части, но и на прилегающих равнинных областях Предуралья и Зауралья в ледниковые эпохи даже в летние месяцы не стаивали и покрывали обширные территории междуречий и речных долин (Антропоген …, 1965; Кайнозой …, 1970), что также способствовало образованию перигляциальных ландшафтов. Первоначально сухие теплые степи сменились холодными перигляциальными степями, где получила развитие вечная мерзлота (Марков и др., 1965). Очень характерно для перигляциальной фауны отсутствие животных, ведущих подземный, роющий образ жизни, что непосредственно обусловлено широким развитием вечной мерзлоты (Вангенгейм, 1975). Климат межледниковий в целом был относительно теплым и влажным. Природные условия микулинского межледниковья Башкирского Предуралья оказались благоприятными для расширения ареалов видов, тяготеющих к лесным местообитаниям.
На протяжении всего позднего плейстоцена на Южном Урале господствовала травянистая степная и лугово-степная растительность. В среднем плейстоцене выпадают из флоры и последние теплолюбивые представители собственно плиоцена, а с одинцовского времени нет в ней и видов, несвойственных ныне существующим местным флорам Южного Урала (Чигуряева, Яхимович, 1965).
Для предбореала характерно широкое распространение березовых лесов с небольшой примесью хвойных и широколиственных, на юге распространены разнотравно-полынные степи. В бореале роль хвойных, особенно сосен, в лесных массивах возросла. Потепление и уменьшение континентальности климата в раннем голоцене способствовало формированию сомкнутого лесного покрова, хотя в целом для голоцена, как и для оптимумов межледниковий, на территории Предуралья отмечается значительно меньшая роль лесов широколиственных пород по сравнению с Русской равниной (Немкова, 1981; 1992). Во время климатического оптимума (атлантак) в среднегорной полосе Южного Урала широко распространились сосновые леса; климат был теплее современного, в лесах увеличилось количество елей и широколиственных пород. Для суббореала установлено широкое развитие липовых лесов, южнее сменяющихся березовыми лесами с заметной примесью широко-лиственных. В субатлантике распространение типов растительности не отличалось от современного: елово-сосновые леса с примесью липы, дуба и вяза. В самом конце субатлантика на юге Предуралья отмечено увеличение безлесных пространств. В целом в течение голоцена произошла смена растительности от холодных лесостепей до теплых лесостепей с ксерофитами.
Таким образом, на протяжении кайнозоя на Южном Урале дважды произошла смена флор: древняя полтавская вечнозеленая флора в олигоцене сменилась тургайской листопадной широколиственной; в плиоцене сменилась на современную (Яхимович, 1992). Полностью современная флора на территории Южного Предуралья существует с апшерона, хотя не исключено, что она сложилась еще в конце позднего акчагыла (Немкова, 1981; Яхимович, 1992) – то есть еще в доледниковое время.
ПАЛЕОНТОЛОГИЧЕСКИЙ МАТЕРИАЛ
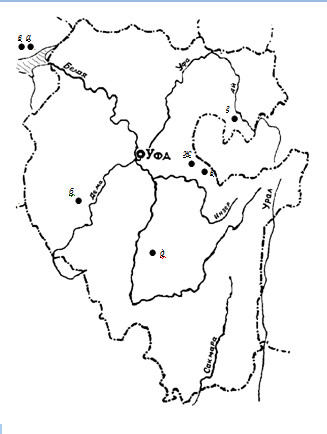
Палеонтологический материал по ископаемым кайнозойским пресмыкающимся непосредственно с территории Южного Урала немногочислен; известные нам местонахождения представлены на схематической карте (рисунок).
Обнаруженные на Южном Урале концентрированные костные остатки ископаемых кайнозойских рептилий (преимущественно в пещерах) своим формированием обязаны, как правило, деятельности хищных птиц, погадки которых «… буквально переполнены остеодермами (щитками) ящериц, косточками лягушек и прочими мельчайшими неопределимыми обломками костей» (Сухов, 1972: с. 137).
Первые сведения относятся к XVIII в. Капитан Николай Рычков упоминает о находках ископаемых позвоночных (не определенных специалистами и, по-видимому, не сохранившихся) в Палянском руднике Берсутского завода близ впадения Белой в Каму (рисунок, а): «Нередко в сих песчаных слоях находят в камень превращенныя рыбы, и виды окаменелых змей … которые … бывают толщиной в обыкновенную палку, и все части покрыты такой же чешуею, как и находимыя тут рыбы» (Рычков, 1770: с. 186-187).
Остальные находки относятся ко второй половине XX в. В отложениях Аккулаевского местонахождения в Давлекановском р-не Республики Баш-кортостан (рисунок, б) обнаружены кости ящериц (Lacerta sp.), датированные эоплейстоценом: в демском горизонте – нижний апшерон – 5 костей, в давлекановском горизонте – средний апшерон – 1 кость (Сухов, 1972). Близ северо-западной границы Башкирии, у дер. Красный Бор из позднеплейстоценовых (микулинское межледниковье; возраст переопределен Яковлевым, 1996) отложений обнаружены (Сухов, 1972а; Чхиквадзе, Сухов, 1977) костные остатки четырех видов рептилий (рисунок, в). Позднеплейстоценовые остатки не определенных даже до семейства змей и ящериц (попадаются лишь отдельные гнезда с костями) известны (Сухов, 1978) из пещеры Кызырбак Салаватского р-на РБ (рисунок, г); позднеголоценовые – из пещеры Зиганская Ишимбайского р-на РБ (рисунок, д). Голоценовые остатки трех видов рептилий известны (Сатаев, Макарова, 1997) из пещеры Заповедная Белорецкого р-на РБ (рисунок, е); позднеголоценовые (возраст – 12800 лет) из отложений пещеры Лемеза IV (рисунок, ж) Архангельского р-на РБ (материалы Т.И.Яковлевой).
Болотная черепаха, Emys orbicularis (L. 1758), из семейства Emydidae – пресноводные черепахи – сформировалась (Хозацкий, 1958) во второй половине плиоцена (эоплейстоцен) в Причерноморье. Ископаемые остатки E.orbicularis встречаются в больших количествах в плейстоценовых и голоценовых отложениях Западной и Центральной Европы, Украины, Кавказа и центральных районов Европейской России, т.е. в основном в пределах границ современного ареала вида (Хозацкий, 1958; Барышников, Несов, 1999). Возможно, на Южном Урале черепаха впервые появились в раннем плейстоцене, хотя в Башкирии ископаемых остатков E.orbicularis пока не обнаружено.
Современная фауна ящериц РБ представлена тремя видами, относящимся к двум семействам: веретенициевые Anguidae и настоящие ящерицы Lacertidae.
Веретеница ломкая, Anguis fragilis L. 1758, известна с миоцена Западной Европы (Основы палеонтологии, 1964). Обычна в четвертичных отложениях европейской части бывшего СССР (Хозацкий, 1982; Зерова, Чхиквадзе, 1984; Ратников, 1988). На Южном Урале известна из позднеплейстоценовых отложений у дер. Красный Бор (Сухов, 1972а; Чхиквадзе, Сухов, 1977), голоценовых отложений пещеры Заповедная (Сатаев, Макарова, 1997), позднеголоценовых отложений пещеры Лемеза IV.
Наиболее древние находки рода Lacerta известны из эоцена Франции и плиоцена Польши и Украины (Основы палеонтологии, 1964), в бывшем СССР – из миоцена Кавказа, нижнего плейстоцена Нижегородской области (Ратников, 1998), плейстоцена Белгородской обл. (Ратников, 1988), а в плиоцене лацертиды были широко распространены в Северном Причерноморье (Чхиквадзе и др., 1983; Зерова, Чхиквадзе, 1984). В Башкирии костные остатки не определенных до вида лацертид – Lacerta sp. – обнаружены в отложениях Аккулаевского местонахождения (Сухов, 1972); возможно, это остатки прыткой ящерицы, Lacerta agilis L. 1758.
L.agilis, как вид, возникла на границе миоцена и плиоцена в пределах со-временного Кавказского перешейка (Прыткая ящерица, 1976). В Казахстане ископаемые находки представителей рода Lacerta не подтвердились (Чхиквадзе и др., 1983). Это может говорить о том, что лацертиды, в частности L.agilis и живородящая ящерица, Lacerta vivipara Jacquin 1787, современный ареал которых включает в себя и северный Казахстан, расселились на его территорию только в послеледниковое время; либо были немногочисленными и не представлены пока в исследованном материале. В ископаемом состоянии на Южном Урале прыткая ящерица обнаружена (Сухов, 1972а; Чхиквадзе, Сухов, 1977) лишь в раскопках у дер. Красный Бор (поздний плейстоцен). Ископаемые остатки L.vivipara известны только из голоценовых отложений пещеры Заповедная (Сатаев, Макарова, 1997) и позднеголоценовых отложений пещеры Лемеза IV.
Змеи, обитающие на территории Башкирии, относятся к двум семействам: ужеобразные Colubridae и гадюки Viperidae.
Colubridae известны из четвертичных отложений Молдавии, Украины и европейской части России (Хозацкий, 1982; Зерова, Чхиквадзе, 1984). Остатки змей из рода Elaphe обнаружены в эоплейстоценовых отложениях Польши (Хозацкий, 1982). Остатки узорчатого полоза, Elaphe dione (Pallas 1773), известны из плейстоцена Крыма (Зерова, Чхиквадзе, 1984), а также из нижнего плейстоцена Нижегородской обл. (Ратников, 1998), т.е. гораздо севернее современного ареала этого вида. Ископаемые остатки водяного ужа, Natrix tessellata (Laurenti 1768), известны из плейстоцена Крыма (Зерова, Чхиквадзе, 1984), плейстоцена Белгородской обл. (Ратников, 1988). Остатки обыкновенного ужа, Natrix natrix L. 1758, обнаружены в отложениях среднего плейстоцена Грузии (Барышников, Несов, 1999), Украины (Зерова, Чхиквадзе, 1984), нижнего плейстоцена Нижегородской обл. (Ратников, 1998). Близ северо-западной границы Башкирии, у дер. Красный Бор (Сухов, 1972а; Чхиквадзе, Сухов, 1977) обнаружены позднеплейстоценовые костные остатки N.natrix. Этот же вид известен из позднеголоценовых отложений пещеры Лемеза IV. Ископаемые остатки обыкновенной медянки, Coronella austriaca Laurenti 1768, а также E.dione, N.tessellata на территории Башкирии на сегодняшний день не обнаружены.
Древнейшие представители семейства Viperidae известны с миоцена (Чхиквадзе и др., 1983); остатки виперид также обнаружены в эоплейстоцене Польши (Хозацкий, 1982), нижнем плейстоцене Нижегородской обл. (Ратников, 1998). Предполагается, что наиболее древней формой гадюк Евразии является степная гадюка, Vipera ursini (Bonaparte 1835), ареал которой сформировался в степях и пустынях Западной Азии (Гаранин, 1983). Обыкновенная гадюка, Vipera berus (L. 1758), сформировалась, по-видимому, только в конце ледникового периода в Центральной Европе (Никольский, 1947).
Ископаемые остатки гадюки обыкновенной известны из антропогена Украины (Зерова, Чхиквадзе, 1984), на Южном Урале – из голоценовых отложений пещеры Заповедная (Сатаев, Макарова, 1997) и позднеголоценовых отложений пещеры Лемеза IV. Костные останки предположительно V.berus, датированные поздним плейстоценом, обнаружены у дер. Красный Бор (Сухов, 1972а; Чхиквадзе, Сухов, 1977). Ископаемые остатки степной гадюки, V.ursini , в Башкирии на сегодняшний день не обнаружены.
ЭТАПЫ ФОРМИРОВАНИЯ СОВРЕМЕННОЙ ФАУНЫ ПРЕСМЫКАЮ-ЩИХСЯ БАШКИРИИ
В палеогене уже существовали все семейства пресмыкающихся фауны бывшего СССР, дожившие до современности (Бакрадзе, Чхиквадзе, 1988). Среднеолигоценовое похолодание не могло не сказаться на видовом составе и распространении пресмыкающихся: очевидно, уже тогда на всей территории бывшего СССР произошло сокращение ареалов влаголюбивых и теплолюбивых групп рептилий. Предполагается, что именно в конце среднего эоцена и начале среднего олигоцена произошли наиболее значительные изменения в составе герпетофауны Казахстана, Кавказа и Западной Европы (Бакрадзе, Чхиквадзе, 1988).
Если современная герпетофауна Средней Азии и Казахстана начала формироваться не позднее среднего олигоцена – раннего миоцена (Чхиквадзе и др., 1983), то для Южного Урала, как и в целом для бывшего СССР (Зерова, Чхиквадзе, 1984), эту границу следует перенести предположительно на конец плиоцена. Все роды и большинство рецентных видов рептилий, составляющих современную фауну РБ, сформировались уже к началу плиоцена. В плиоценовых отложениях Башкирии обнаружены пока только ископаемые Lacertidae, но, вероятно, будут найдены и представители по крайней мере еще трех семейств: Anguidae, Colubridae, Viperidae. Пока не известны (не определены?) ископаемые остатки плиоцен-голоценовых пресмыкающихся, относящихся к родам и видам, отличным от современных; также нет пока данных об ископаемых находках современных видов, не обитающих ныне на Южном Урале, но, возможно, обитавших там ранее (например, возможное обитание в интерстадиалах позднего плейстоцена либо в период голоценового климатического оптимума на территории Южного Урала обыкновенного щитомордника, Agkistrodon halys (Pall. 1775), в настоящее время отсутствующего даже на юге Оренбургской обл.). Скудность палеонтологического материала о герпетофауне кайнозоя Южного Урала не позволяет сделать более точные выводы и заключения.
Похолодания плиоцен – эоплейстоцена и оледенения плейстоцена оказали огромное влияние на фауну пресмыкающихся Южного Урала (как и Палеарктики в целом). Хотя покровные оледенения не распространялись на территорию Башкирии за исключением, возможно, окского оледенения (Монин, Шишков, 1979), тем не менее перигляциальная зона холода и мерзлотные явления охватывали весь Уральский регион (Величко, 1975), южная граница распространения вечной мерзлоты в конце валдайского оледенения доходила до 46-48° с. ш., а районы древнего сезонного промерзания сибирского типа достигали Кавказа и северного Прикаспия включительно (Палеогеография Европы …, 1982), т.е. мерзлотой оказалась охвачена по существу вся Восточно-Европейская равнина (Величко, 1975) и значительная часть Западно-Сибирской равнины. На территории европейской части России сформировался сложный комплекс криогенных образований, ведущую роль в котором играли крупные трещинные системы размером в среднем 20-30 м при средней толщине трещин 4-5 м (Величко, 1975). К востоку от Уральского хребта, на территории Южного Зауралья в это время «… часто наблюдаются криогенные деформации (морозобойные клинья, инволюции), указывающие на длительное промерзание грунтов и суровый холодный климат» (Стефановский, Малеева, 1977: с. 52). С другой стороны, хотя в горных районах Южного Урала даже в самые холодные периоды находились местообитания, где произрастала лесная растительность, в западных предгорьях сохранялись и представители широколиственной флоры (Историческая экология животных гор …, 1990), а в зоне развития сезонной мерзлоты обнаружены ископаемые остатки ведущего подземный образ жизни цокора (Мекаев, 1987), тем не менее для пойкилотермных животных климатические условия были слишком суровыми и на большей части Русской равнины, Урала и Сибири (Никольский, 1947) пресмыкающиеся вымерли. Возможно, именно в это время рептилии умеренного пояса окончательно сформировали способность впадать в зимнюю спячку, как это предполагается (Мензбир, 1934) для грызунов потретичной фауны.
Неоднократные наступления и отступления ледников привели к тому, что ареалы видов периодически сжимались, отступая в периоды похолоданий в рефугиумы, и расширялись в периоды потеплений. Сам факт обнаружения ископаемых пресмыкающихся на Южном Урале в местонахождениях плейстоценовых межледниковий (Сухов, 1972а; Чхиквадзе, Сухов, 1977; Сухов, 1978) позволяет говорить об удаленности ледников и относительно теплом климате данного региона в периоды интерстадиалов (Ратников, 1996). Число возможных рефугиумов спорно: на материале по водоплавающим птицам Кривенко (1991) выделяет 10 центров расселения на территории бывшего СССР: Восточно-Тянь-Шаньский, Копетдагский, Западно-Казахстанский и др. По мнению Никольского (1947), убежища были только на западе Франции и в Маньчжурии, а рефугиумы, выделяемые для птиц (Кривенко, 1991) в азиатской части Евразии, видимо, не были пригодны для обитания рептилий (Никольский, 1947). Для герпетофауны Уральского региона наибольшее значение имели Балканский, Кавказско-Малоазийский и, возможно, Средне-Азиатский ледниковые убежища, их которых в дальнейшем началось вторичное распространение пресмыкающихся в границы современного ареала. Расселение животных из рефугиумов началось по мере постепенного потепления климата, начавшегося 12-8 тыс. лет назад в предбореальный период голоцена (Кривенко, 1991). Расширение ареалов активизировалось в период голоценового климатического оптимума. Основное направление расселения пресмыкающихся, входящих в состав фауны РБ, происходило из рефугиумов южной и юго-восточной Европы, Кавказа на север и северо-восток, через Уральские горы, что подтверждается данными о клинальной изменчивости некоторых морфологических, фенетических и других признаков (Прыткая ящерица, 1976; Анисимова, 1981). В суббореальный и субатлантический периоды голоцена после некоторого похолодания и последовавшего за ним сокращения ареалов животных климатические условия стабилизировались (Кривенко, 1991).
Тот факт, что рассматриваемые нами виды имеют разный «исторический возраст», мало отразился на современной картине их ареалов. После окончания последнего оледенения у всех видов были примерно одинаковые стартовые условия при расселении с той лишь разницей, что одни виды начали свое послеледниковое расселение с запада из рефугиумов Центральной и Южной Европы (E.orbicularis, A.fragilis, L.agilis, L.vivipara, N.natrix, V.berus), другие – с юга, из Западной Азии (V.ursini и, возможно, N.tessellata), третьи, возможно, – с Дальнего Востока (E.dione).
Различия в площади и границах современных ареалов можно попытаться объяснить разной скоростью расселения и/или различной адаптивностью различных видов. Молодые в эволюционном смысле виды, прежде всего живородящая ящерица и обыкновенная гадюка, сформировались во многом под влиянием перигляциальных условий и поэтому, во-первых, более приспособлены к условиям умеренного климата, во-вторых, более экологически пластичны. Эти качества позволили им очень быстро в течение позднеплейстоцен-голоценового времени распространиться на огромные территории на восток через всю Сибирь, на север за Полярный круг. Обыкновенные уж и прыткая ящерица имеют достаточно пространный современный ареал, как и узорчатый полоз, хотя последний расселялся с востока. Обыкновенная ме-дянка, веретеница ломкая и отчасти болотная черепаха, расселяясь из Европы на восток, имеют сопоставимые современные ареалы, захватывающие небольшую часть Западно-Сибирской равнины, т.е. все эти три вида лишь недавно перешли через Уральские горы.
Для Южного Урала полностью современная герпетофауна начала формироваться только в послеледниковое время при расселении пресмыкающихся из ледниковых рефугиумов, т.е. полностью состоит из позднечетвертичных мигрантов. С этих позиций вопрос формирования современной фауны пресмыкающихся сводится к изучению характера их послеледникового расселения. Видовой состав герпетофауны РБ относительно беден; нет эндемиков, что неудивительно, учитывая суровые перигляциальные климатические условия антропогена. Современный набор видов существовал, по крайней мере в Предуралье, уже в среднем голоцене, в период голоценового климатического оптимума. Дальнейшие исследования, несомненно, позволят уточнить хронологию и детали проникновения современных видов на территорию Башкирии в ходе плейстоцен – голоценового расселения.
ВЛИЯНИЕ УРАЛЬСКИХ ГОР И АНТРОПОГЕННЫХ ФАКТОРОВ НА ФОРМИРОВАНИЕ ФАУНЫ ПРЕСМЫКАЮЩИХСЯ БАШКИРИИ
Южный Урал входит в пределы Урало-Сибирского пояса каледонской и герцинской складчатости, завершившего геосинклинальный этап развития в конце палеозоя (Башенина, 1948; Монин, Шишков, 1979). До конца олигоцена горы Южного Урала были низкими: их абсолютные высоты в среднем составляли всего 300-500 м, наивысших за весь кайнозой высот достигли в миоцене (Яхимович, 1992), в дальнейшем лишь незначительно сгладились. В целом рельеф Южного Урала образован слабодифференцированными сводово-глыбовыми подвижками неоген-четвертичного времени.
Из десяти видов герпетофауны РБ «стандартные» шесть видов рептилий – три вида ящериц (A.fragilis, L.agilis и L.vivipara) и три вида змей (C.austriaca, N.natrix и V.berus) распространены на всей территории Башкирии, относительно хорошо приспособлены к существованию в холодном климатическом поясе и составляют основу современной постгляциальной герпетофауны умеренных широт европейской части России и Западной Сибири. При сравнении фауны пресмыкающихся РБ и сопредельных регионов видно, что герпетофауна северной части Южного Урала идентична таковой Среднего и южной части Северного Урала. Оставшиеся четыре вида (E.orbicularis, E.dione, N.tessellata и V.ursini) обитают лишь в южных районах Башкирии. Южные отроги Уральских гор как бы прогибают северные границы ареалов этих видов к югу (Хабибуллин, 1999).
В настоящее время Уральский хребет не является меридиональной границей ни для одного вида рептилий, обитающих на Южном Урале, т.е. все виды, присутствующие к западу от хребта, обитают и к востоку от него (Хабибуллин, 1999). По всей видимости, горы как физическая преграда в историческом плане существенно не влияли на вектор и скорость распространения пресмыкающихся на Южном Урале, но как ландшафтообразующий фактор их роль значительна и несомненна. Так, наибольшая залесенность характерна для горных районов, а степи, развитые преимущественно в равнинной части республики, в значительной степени распаханы. Так, до начала интенсивного освоения территории Башкортостана степи занимали не менее 1/3 ее равнинной части. После освоения практически всех пахотопригодных земель в Предуралье степи почти исчезли уже в начале этого столетия, а в Башкирском Зауралье – к началу 50-х гг. Вклинивание Уральских гор (и, соответственно, лесных массивов) в лесостепную зону являлось и является благоприятным моментом для распространения на юг бореальных видов. Но, по-видимому, эта «лесопродвигающая» функция Уральских гор возникла относительно недавно и связана с тем, что на значительных площадях в равнинной части республики леса были сведены уже в историческое время. Уральские горы являются водоразделом бассейнов рек Волги, Урала и Оби и в этом плане имеют значение для распространения, а в историческом плане, и для послеледникового расселения связанных с приводными биотопами пресмыкающихся (например, расселение с низовьев Волги и Урала болотной черепахи и водяного ужа).
В заключение добавим, что современные ареалы пресмыкающихся еще окончательно не сложились и часть видов (A.fragilis, C.austriaca) продолжает медленно увеличивать свой ареал на восток. В последние столетия на состояние животного мира большое воздействие оказывает хозяйственная деятельность человека, что в первую очередь проявляется в трансформации природных ландшафтов и сокращении пригодных для обитания участков; однако несмотря на всю свою масштабность, в историческом плане именно на процесс формирования фауны (фауны как набора видов) пресмыкающихся Южного Урала антропогенные изменения существенно влияния не оказывали.
СПИСОК ЛИТЕРАТУРЫ
Анисимова Е.В., 1981. О географической изменчивости полового диморфизма медянки // Вопр. герпетологии. Л.:Наука. С. 9.
Антропоген Южного Урала, 1965. М.: Наука. 272 с.
Бакрадзе М.А., Чхиквадзе В.М., 1988. Материалы к третичной истории герпетофауны Кавказа и сопредельных регионов // Вестник гос. Музея Грузии. XXXIV-A. С. 176-193.
Барышников Г.Ф., Несов Л.А., 1999. О находке болотной черепахи (Emys orbicularis) в ашельской фауне пещеры Кударо 3 в Закавказье // Материалы по истории фауны Евразии. Тр. ЗИН РАН. Т. 277. С. 127-129.
Башенина Н.В., 1948. Происхождение рельефа Южного Урала. М.: ГИГЛ. 232 с.
Вангенгейм Э.А., 1975. О перигляциальной фауне плейстоцена // Палеогео-графия и перигляциальные явления плейстоцена. М.: Наука. С. 83-88.
Величко А.А., 1975. Парагенезис криогенной (перигляциальной) зоны // Палеогеография и перигляциальные явления плейстоцена. М.: Наука. С. 89-100.
Гаранин В.И., 1983. Земноводные и пресмыкающиеся Волжско – Камского края. М.: Наука. 175 с.
Зерова Г.А., Чхиквадзе В.М., 1984. Обзор кайнозойских ящериц и змей СССР // Изв. АН ГССР. Сер. биол. Т. 10. № 5. С. 319-325.
Историческая экология животных гор Южного Урала, 1990. Свердловск: УрО АН СССР. 248 с.
Кайнозой Башкирского Предуралья, 1970. Т. 2. Ч. 3. Этапы геологического развития Башкирского Предуралья в кайнозое. М.: Наука. 136 с.
Кривенко В.Г., 1991. Водоплавающие птицы и их охрана. М.: Агропромиздат. 271 с.
Марков К.К., Лазуков Г.И., Николаев В.А., 1965. Четвертичный период (ледниковый период – антропогеновый период) Т. 2. Территория СССР. М.: Изд-во МГУ. 436 с.
Мекаев Ю.А., 1987. Зоогеографические комплексы Евразии. Л.: Наука. 126 с.
Мензбир М.А., 1934. Очерк истории фауны Европейской части СССР. М.: Биомедгиз. 224 с.
Монин А.С., Шишков Ю.А., 1979. История климата. Л.: Гидрометеоиздат. 408 с.
Немкова В.К., 1981. Флоры и растительность Предуралья в плиоцене, плейстоцене и голоцене // Плиоцен и плейстоцен Волго-Уральской области. М.: Наука. С. 69-77. – 1992. Флоры и растительность Предуралья в плиоцене, плейстоцене и голоцене // Флора и фауна кайнозоя Предуралья и некоторые аспекты магнитостратиграфии. Уфа: БНЦ УрО РАН. С. 11-32.
Никольский А.М., 1947. Роль ледникового периода в истории фауны палеарктической области // Бюл. Моск. о-ва испытат. природы. Отд. биол. Т. 52. Вып. 5. С. 3-14.
Основы палеонтологии, 1964. Земноводные, пресмыкающиеся, птицы / Под ред. Рождественского А.Н., Татаринова Л.П. М.: Наука. 722 с.
Палеогеография Европы за последние сто тысяч лет (атлас-монография), 1982. М.: Наука. 156 с.
Прыткая ящерица, 1976. М.: Наука. 376 с.
Ратников В.Ю., 1988. Верхнечетвертичные герпетофауны Белгородской области // Палеонтол. журн. № 3. С. 119-122. – 1996. К методике палеогеографических реконструкций по ископаемым остаткам амфибий и рептилий позднего кайнозоя Восточно-Европейской платформы // Палеонтол. журн. № 1. С. 77-83. – 1998. Остатки пресмыкающихся из нижнеплейстоценового местонахождения Березовка Нижегородской области // Палеонтол журн. № 3. С. 74-76.
Рычков Н.П., 1770. Журнал, или дневные записки путешествия Капитана Рычкова по разным провинциям Российского государства, 1769 и 1770 году. С-Пб. 190 с.
Сатаев Р.М., Макарова О.В., 1997. Ископаемая герпетофауна из пещеры “Заповедная” // Ежегодник-96. Уфа: ИГ УНЦ РАН. С. 14-15.
Синицын В.М., 1980. Природные условия и климаты территории СССР в раннем и среднем кайнозое. Л.: Изд-во ЛГУ. 104 с.
Стефановский В.В., Малеева А.Г., 1977. Палеогеографическая обстановка ранней половины позднеплейстоценового времени на территории Южного Зауралья // Фауна Урала и Европейского Севера. Свердловск: Изд-во УрГУ. Сб. 5. С. 31-34.
Сухов В.П., 1972. Позвоночные – Vertebrata (мелкие) // Фауна и флора Аккулаева (опорный разрез среднего акчагыла – среднего апшерона Башкирии). Уфа: Изд-во БашФАН СССР. С. 119-139. – 1972а. О находке остатков среднеплейстоценовых мелких позвоночных у дер. Красный Бор на р. Каме // Вопросы стратиграфии и корреляции плиоценовых и плейстоценовых отложений северной и южной частей Предуралья. Вып. 1. Уфа. С. 133-136. – 1978. Позднеплейстоценовые и голоценовые мелкие позвоночные из пещер западного склона Южного Урала // К истории позднего плейстоцена и голоцена Южного Урала и Предуралья. Уфа. С. 64-85.
Хабибуллин В.Ф., 1999. О границах ареалов пресмыкающихся на Южном Урале // Изучение и охрана биологического разнообразия ландшафтов Русской равнины: Сб. мат-лов. Пенза. С. 286-287. – 1999а. Пресмыкающиеся Республики Башкортостан // Автореф. дис. … канд. биол. наук. – Уфа: Изд-во БашГУ. 18 с.
Хозацкий Л.И., 1958. Современное и прошлое распространение черепах в СССР // Проблемы зоогеографии суши: Мат-лы совещ. Львов. С. 319-324. – 1982. Пресмыкающиеся // Стратиграфия СССР. Четвертичная система. Полутом 1. М.: Недра. С. 252-262.
Чигуряева А.А., Яхимович В.Л., 1965. О флоре и растительности Южного Приуралья в среднеакчагыльское-голоценовое время // Антропоген Южного Урала. М.: Наука. С. 164-187.
Чхиквадзе В.М., Сухов В.П., 1977. Земноводные и пресмыкающиеся из чет-вертичных отложений Красного Бора (р. Кама) // Вопр. герпетологии. Л.: Наука. С. 227-228.
Чхиквадзе В.М., Шаммаков С.Ш., Зерова Г.А., 1983. К истории формирования фауны чешуйчатых рептилий (Squamata) Средней Азии и Казахстана // Изв. АН Туркм. ССР. Сер. биол. наук. № 2. С. 3-8.
Яковлев А.Г., 1996. Мелкие млекопитающие плейстоцена и голоцена Башкирского Предуралья и западного склона Южного Урала // Автореф. дис. … канд. биол. наук. Екатеринбург: ИЭРиЖ УрО РАН. 17 с.
Яхимович В.Л., 1992. Смена состава флор, ландшафтов и климатические изменения в палеогене и неогене Предуралья // Флора и фауна кайнозоя Предуралья и некоторые аспекты магнитостратиграфии. Уфа: БНЦ УрО РАН. С. 4-10.
Добавить комментарий