Хабибуллин А.Ф., Хабибуллин В.Ф.
Зоол. Журн. 2012. Т. 91, № 3. – С. 304-309.
В городской среде человек посредством агротехнических мероприятий, в частности, формовочной обрезки, влияет на санитарное и декоративное состояние городских зеленых насаждений. Показана неоднозначность регуляторной деятельности человека для отдельных взаимосвязанных членов триотрофа «растение – энтомофитофаг – энтомофаг». Отмечено преобладание «плюсов» агротехнических мероприятий для человека и растения, «минусов» – для энтомофитофага и энтомофага. Динамика таких «нарушений» в пространстве и времени обуславливает существование микросукцессий заселения растений фитофагами и энтомофагами.
Ключевые слова: урбоэкосистема, триотроф «растение – энтомофи-тофаг – энтомофаг», городские насаждения, шиповник, большая розанная тля, Adalia bipunctata.
ВВЕДЕНИЕ
В условиях глобальной антропогенной трансформации среды значительное внимание по праву уделяется экологическим проблемам горо-да.
Весомым фактором улучшения экологической ситуации в городах является озеленение. Зеленые насаждения являются органической частью планировочной структуры современного города; при этом большое значение придается декоративности и эстетическому состоянию насаждений. Озелененные территории городов и пригородных зон оказывают сущест-венное влияние на многие показатели качества окружающей среды, имеют оздоровительное, средозащитное и средоформирующее значение. Экологическая эффективность зеленого фонда городов зависит от двух главных факторов – их площади и биологического разнообразия, в первую очередь – растительного мира, предопределяющего разнообразие гетеротрофной биоты (Едренкина, 2005).
Видное место в животном населении городских экосистемах занимают энтомокомплексы, в трофической структуре которых преобладают фитофаги (Киселев, 2005). Город представляет собой специфическую для насекомых-фитофагов среду обитания. С одной стороны, снижение уровня антибиоза кормовых растений под влиянием урбогенных ноксов, уменьшение пресса паразитов и хищников способствуют росту численности популяций энтомофитофагов. С другой стороны, на их численность отрицательно влияет «островная» пространственная структура зеленых насаждений городов, прямое и косвенное (через кормовое растение) воздействие поллютантов (Тарасова, 2004).
Особую опасность для древесных и кустарниковых насаждений города представляют насекомые – филлофаги; в первую очередь это касается тлей (Aphidoidea), партеногенез и ускоренное развитие которых обеспечивает быструю и массовую колонизацию кормовых ресурсов и их стремительную утилизацию (Дьяконов, 2003). Афидофауны отдельных российских городов насчитывают десятки видов, например, в Воронеже – 68 (Кузьминов, 2005). Тли, как правило, живут колониями на нижней стороне листьев, на молодых побегах, цветоножках, питаясь соками растений. Они представляют опасность тем, что ослабляют растение, снижают его устойчивость к болезням, а также могут являться переносчиками вирусных заболеваний (Щербакова, Карпун, 2008). Компактно растущие городские моновидовые посадки кустарников особенно подвержены поражению быстро размножающимися тлями.
Определенное значение для биоты города имеют загрязнения, в первую очередь атмосферные (Тарасова, 2004). Так, покрывающая сплошным слоем поверхность листьев пыль делает их малопривлекательными для насекомых с ротовым аппаратом грызущего, но не колюще-сосущего типа (Баранник, 1979); последние в этом плане особенно устойчивы к аэрозагрязнениям – как промышленным, так и автотранспортным. Поэтому плотность населения тлей на уличных деревьях во много раз больше, чем на деревьях за городом или в крупных лесопарках; в свою очередь, увеличивается численность хищников и паразитов тлей. Однако, наличие сосущего ротового аппарата вынуждает тлей питаться на сочных частях растений с тонкими покровами; и в процессе роста и развития кормового растения тли вынуждены перемещаться на более молодые листья верхнего яруса, боковых и порослевых побегов. Сильно одеревеневшие, с грубыми покровами ветви тли не заселяют; уязвимы лишь молодые побеги, вегетирующие в текущем году.
В озеленении улиц, городов, садов и парков широко применяют различные виды шиповника; как наиболее газоустойчивые упоминаются ши-повники роза морщинистая Rosa rugosa Thunberg 1784, и роза даурская Rosa davurica Pallas 1789 (Зей-Нечаева, 1984). Тли являются обычными вредителями шиповника в городе (Колесников, Болдырев, 2007). Так, энтомофауна шиповника в искусственных биогеоценозах г.Уфы насчитывает 21 вид: 17 энтомофитофага и 4 энтомофага (Зей-Нечаева, 1984). Для Европейской части России к группе факультативно опасных отнесены большая розанная тля Macrosiphum rosae (L. 1758), розанная листовая тля Capitophorus tetrarhodus (Walker 1849). Считается, что вредоносность этих видов носит очаговый характер и в целом не превышает 5% (Колесников, Болдырев, 2007).
Большинство работ, посвященных изучению системы триотрофа «растение – фитофаг – энтомофаг» направлено на биохимические составляющие взаимосвязей (влияние метаболитов кормового растения на фитофагов и через них – на паразитов) (Мартемьянов, Бахвалов, 2007). Гораздо меньшее внимание уделено агротехническим методам (в широком смысле – применительно не только к почвам, но и к растительности, животному населению), несмотря на их важную роль в системе мероприятий по защите зеленых насаждений. Так, практически все виды тлей зимуют на побегах, в трещинах коры, откуда они весной заселяют молодые вегетирующие листья: в этом аспекте старые, непрореженные посадки с отсыхающими ветками являются источником распространения тлей, а периодическая обрезка таких веток подрывает кормовую базу тлей. В некоторых случаях вспышки численности вредителей городских зеленых насаждений тесно связаны именно с ошибками в технологии и сроках возделывания и защиты растений, а также с пренебрежением проведения санитарных и профилактических работ. Регуляция численности ряда групп насекомых может осуществляться путем целого спектра мероприятий: периодической обрезки деревьев и кустарников, омоложения старых посадок, обработкой почвенного покрова и др. Работы по уходу за зелеными насаждениями сказываются не только на растениях, но и на биоценотически (трофически, топически и пр.) связанные с ними группы организмов.
В данной работе мы рассмотрим влияние проводимых в городе формовочных агротехнических мероприятий на триотроф «растение – энтомофитофаг – энтомофаг».
МАТЕРИАЛ И МЕТОДЫ
Изучение влияния человека на систему (триотроф) «растение – фитофаг (энтомофитофаг) – энтомофаг» проведем на примере влияния формовочной обрезки кустарника на фитофагов и их хищников.
Материал собран в городе Уфа (Республика Башкортостан). Фенологические наблюдения над энтомофагом проводились с 2000 года, по системе триотрофа – в 2008-2009 гг. в южной части города. Количественные данные были собраны в июне 2009 года на модельном участке – газоне длиной около 100 м и шириной 8 м между тротуаром и проезжей частью, расположенном на четной стороне улицы Пушкина близ перекрестка с улицей Аксакова.
В качестве автотрофного компонента триотрофа исследовался шиповник роза морщинистая R.rugosa.
Посадки шиповника расположены линейно вдоль тротуара; исходно сплошная лента живой изгороди (ширина 0.8-1.5 м) со временем оказалась фрагментирована промежутками 0.3-1.5 м. Отдельные кусты шиповника имеют трапециевидную форму, плавно расширяясь от 70см у основания до 110-130 см у линии обреза. Ветвление начинается на высоте 50-80 см. Расстояние от основания кустарника до асфальта порядка 20 см.
В качестве энтомофитофага выбрана большая розанная тля M.rosae. В Уфе этот вид, наряду с розанным нисходящим пилильщиком Ardis brunniventris (Hartig 1837), розанной мухой Rhagoletis alternata (Fallen 1814) и листовертками (5 видов) считается наиболее практически значимым (Зей-Нечаева, 1984).
В качестве энтомофага изучалась двуточечная божья коровка Adalia bi-punctata (L. 1758) (Coleoptera: Coccinellidae). Это самый массовый из отмеченных нами на шиповнике 10 видов кокцинеллид. Встречающиеся имаго – в основном типичной формы (typica): красный фон надкрылий с двумя округлыми черными пятнами; черные формы (sexpustulata и quadrimaculata) редки (менее 4%). Личиночные стадии дифференцированы (по Савойской, 1983) на две группы: младшие – 1-2 возраста и старшие – 3-4 возраста.
В первый день наблюдений мы застали собственно сам процесс обрезки шиповника: западная часть участка была уже обработана, восточная – нет. Сначала мы осмотрели не обработанную (интактную) часть посадок кустарника, затем – обработанную (импактную). В последующие дни наблюдения проводились ежедневно. В качестве контроля изучались не подвергаемые формовочной обрезке внутриквартальные насаждения шиповника.
Все количественные данные по насекомым приводятся в расчете на один погонный метр кустарниковой изгороди.
РЕЗУЛЬТАТЫ
Интактные кусты. Фенологический календарь шиповника в условиях Уфы таков: облиствление – вторая декада мая, цветение – третья декада мая – вторая декада июня, начало созревания плодов – третья декада июня.
Энтомофитофаг. 15-25% молодых (не скелетных) побегов шиповника заселено тлями. Начало питания тлей на шиповнике – вторая декада мая, окончание – вторая декада августа. За сезон развивается до 10 генераций.
Энтомофаг. В условиях Южного Урала A.bipunctata бивольтинна; на шиповнике развивается первая генерация. Яйцекладка начинается с первой декады июня, личинки появляются со второй декады июня, имаго второй генерации – в третьей декаде июня.
На модельном участке обнаружены все стадии развития энтомофага A.bipunctata. Имаго – численность 4-8 шт./м. Имаго сосредоточены на молодых побегах; единичные особи в поисковом поведении заходят на скелетные ветви. Яйца – численность 0,6 шт./м. Яйцекладки расположены на нижней стороне листьев молодых побегов. Личинки – численность 4,4 шт./м. Личинки разных возрастов обнаружены только на молодых побегах, но не на скелетных. Около трети обнаруженных личинок поедали тлей, остальные передвигались. Куколки – численность 0,8 шт/м, расположены в дистальной (верхней) трети молодых побегов.
Импактные кусты.
Шиповник относится к кустарникам, возобновление которых после об-резки происходит за счет крупных боковых побегов – стеблевой поросли. Су-ществующие в городе плановые формовочные и санитарные обрезки крон деревьев и кустарников предусматривают в том числе и плоскостную обрезку: сверху на высоте 0.5-1.5 м от поверхности земли, а также с боков, срезаются все вертикальные побеги, придавая кустарнику необходимый поперечный профиль. На наблюдаемом нами участке формовочная обрезка шиповника производится на высоте 1.4-1.6 м. Задача формирования куста – получение крепких, основных скелетных ветвей, на которых в дальнейшем образуются побеги последующих порядков. Практики (см., напр., Серебрякова и др., 1977) настоятельно советуют удаление и немедленное сжигание срезанных ветвей по окончании работ.
В ежегодно подстригаемом кустарнике можно выделить: во-первых, сильно одревесневшие многолетние ветви – назовем их «скелетные» – от земли до уровня плоскости обреза; во-вторых, ежегодно отрастающие после обрезки слабо одревесневающие побеги – назовем их «молодые». Принимая во внимание ежегодный характер обрезки, проводимой в Уфе в середине лета, к молодым можно отнести побеги, выросшие в конце предыдущего теплого сезона (июль-сентябрь) и в начале текущего (май-июнь). Обрезаются побеги длиной 20-50 см, остаются только скелетные ветви, без молодых побегов и листьев. Сохраняются единичные побеги – по периферии куста (не доросли до плоскости среза), а также выше плоскости среза (некачественная обрезка): такие пропущенные при обрезке побеги мы далее будем называть соответственно «боковыми» и «случайными» (рис.). Отстриженные побеги складываются у основания кустарника, по обеим сторонам живой изгороди, в том числе на асфальт (до 30 см вглубь тротуара).
Энтомофитофаг. После обрезки на импактных кустах тлей не обнаружено. Удаление молодых побегов существенно снижает численность оставшихся на шиповнике тлей; возможно, лишь единичные особи могут сохраниться на случайных побегах. Срезанные побеги в жаркую погоду быстро иссыхают. Теоретически за сутки бескрылые тли могут перемещаться от первичного растения на 10–18 м (Дьяконов, 2003), однако в наблюдаемой ситуации большинство тлей продолжало питание не срезанных побегах (до момента их высыхания), и за это время находящиеся на этих же побегах личинки успевали уничтожить тлей. Часть крылатых особей тлей улетает.
Энтомофаг. Имаго – численность 0.1 шт./м. Непосредственно в процессе обрезки кустарника потревоженные жуки падают на землю либо улетают. На уже обработанных кустах обнаружены единичные имаго, большинство же перелетели за пределы обследуемого участка либо (единицы) на временно интактную часть посадок. В последующие дни зафиксированы единичные особи имаго (на весь участок – 2-3 особи).
Яйца – на оставшихся после обрезки скелетных ветвях, как и на «случайных» яйцекладки энтомофага не обнаружены.
Личинки – единичные личинки обнаружены только на боковых и «слу-чайных» побегах – 0.2 шт./м. Часть личинок во время обрезки от меха-нических воздействий рабочих и сотрясения побегов падает на землю; другая часть оказывается на земле вместе со срезанными побегами, не покидая их. За пределами срезанных побегов – на асфальте и траве – личинки не обнаружены. Наблюдения в последующие дни показали, что личинки так и остаются на тех же самых ветках, и, уничтожив доступных тлей, гибнут: сначала личинки младших возрастов, позднее – старших.
Личинки с упавших на асфальт побегов испытывают воздействие высо-ких температур, и проявляют высокую активность, передвигаясь в пределах отстриженного побега. Однако на асфальт с отрезанного побега не переходят (три наблюдения по 15 мин), в том числе при «подталкивании». Двигательная активность непродолжительна, после интенсивных хаотичных перемещений личинки останавливаются, сжимаются и остаются неподвижными (при физическом воздействии – подталкивании – лишь отодвигаются). Гибель у находящихся на упавшем на асфальт побеге личинок наступает через 20-40 мин. У двух личинок старшего возраста, сня-тых с побега и помещенных на открытый асфальт, наблюдали хаотичные аномально активные перемещения. Продолжительность движений до остановки и гибели (вследствие теплового шока: напомним, в период наблюдений температура воздуха превышала 30ºС, температура асфальта – еще выше) составила 25-30 с. В другом наблюдении личинкам старшего возраста с упавших побегов было «предложено» перейти на другой побег (предлагались лист подорожника, сорванный непосредственно у живой изгороди и другие побеги шиповника; субстраты совмещали в одной плос-кости перед личинкой) – перешли 3 из 4 протестированных личинок.
Куколки – после обрезки на импактных кустах куколок не обнаружено.
ОБСУЖДЕНИЕ
Формовочная обрезка кустарника отрицательно влияет на преимагинальные стадии развития кокцинеллид. Особенно катастрофичны последствия для уже отложенных яиц. Так как качество обрезки высокое (в среднем один «случайный» побег на куст), то яйцекладки полностью уничто-жаются: ни на одном из оставшихся не обрезанном отдельном побеге яиц не обнаружено. Если допустить равномерность процесса яйцекладки среди всех молодых побегов, то вероятность выживания даже оставшаяся случайно яйцекладок на таком изолированном побеге, уязвимом воздействию солнца, ветра, механического повреждения и пр., невелика. Вышедшие в таких условиях личинки были бы обречены на голодание (ввиду недостатка тлей) и не смогли бы завершить развитие. Шансов на успешное развитие у яиц, находящихся на упавших в траву побегах, нет. Даже несовершенная технология проведения работ (обрезанные побеги не удаляют – и через две недели после окончания работ обрезки так и лежали близ кустарника) не повышает выживаемость кокцинеллид: гибнут и яйца, и личинки.
Рабочая гипотеза о перемещении (точнее, вползании) личинок со срезанных побегов на кустарник и дальнейшее успешное развитие не под-твердилась. Во-первых, личинки не проявляли активности по перемещению с упавших побегов; во-вторых, личинкам трудно добраться до кустарника и найти подходящие (не скелетные) побеги; в-третьих, личинки неизбежно столкнутся с недостатком пищевых объектов (тлей) на обработанном кустарнике.
Что касается куколок, то теоретически из «зрелых» куколок может вый-ти стадия имаго; но эта возможность стремительно уменьшается по мере нахождения куколок на земле среди увядающих срезанных побегов.
Специалисты признают пользу проведения санитарных и формовочных обрезок, подчеркивая необходимость срезания жировых и порослевых побегов, на которых в большинстве случаев зимуют яйца тли. Более того, во избежание заражения здоровых растений срезанные побеги советуют сжечь до начала выхода из яиц самок-расселительниц (Кузьминов, 2005). И хотя формовочная обрезка положительно влияет на санитарное состояние и внешний вид кустарника, является эффективным приемом регуляции численности тли, ее воздействие на численность энтомофагов следует при-знать отрицательным. Дело не только в уничтожении тлей (все-таки это способствует здоровью кустарника) как кормовой базы для энтомофагов, но в фактически прямом уничтожении яиц, личинок и отчасти куколок.
Неоднозначно можно оценить и влияние агротехнических мероприятий на кормовое растение: чего в проводимой обрезке кустарника больше – плю-сов или минусов? Для шиповника – плюс (избавление от вредителей и омоложение), для тлей – минус (гибель и потеря кормовой базы), для энтомофагов – минус (гибель преимагинальных стадий и потеря кормовой базы), для человека – плюс (здоровая и эстетическая живая изгородь). По последнему пункту хотелось бы подчеркнуть, что нельзя игнорировать интересы горожан: ведь город создан и функционирует в первую очередь как среда обитания человека, и все мероприятия, в том числе по оптимизации и управлению природных элементов, должны быть направлены на создание комфортных и удобных условий для жизни человека. В целом плюсов, на наш взгляд, больше, ибо выполняется первостепенная задача по формированию живой изгороди надлежащего санитарного состояния с высокими декоративными свойствами; попутно регулируется численность вредителей. В иерархии целей: поддержание высокой численности энтомофагов либо обеспечение эстетики и «здоровья» зеленых насаждений – преимущество следует отдать последним.
Попытаемся «оправдать» и невольно наносимый ущерб энтомофагам. Известно, что кокцинеллиды-афидофаги в природе систематически сталкиваются с нестабильностью кормовой базы (Яблоков-Хнзорян, 1976) – ведь их цикл развития недостаточно краток для гибкого следования за колебаниями численности жертвы (так называемая эфемерность колоний тлей), и в естественных условиях часто встречается ситуации, когда часть генерации коровок не успевает благополучно завершить развитие до момента исчезновения пищевых объектов. Поэтому в рассматриваемом нами случае для коровок нет ничего «критического», что косвенно подтверждается многочисленностью в городах представителей этого семейства, и особенно антропофильной двуточечной коровки.
Отмечаемая в некоторых случаях слабая эффективность кокцинеллид как агентов биологического контроля определяется их неспособностью погасить раннюю (майскую) вспышку численности тлей (см., напр., Ченикалова и др., 2008): количество перезимовавших коровок невелико, а их наиболее прожорливая стадия – личинки – еще не успели развиться из откладываемых в конце весны яиц. Онтогенез тлей гораздо лабильнее: цикл развития от яйца до имагинальной фазы при оптимальных условиях заканчивается за 5–7 дней (у двуточечной коровки – около 30, у других больше), а благодаря партеногенетическому размножению афидидами достигается почти 8-кратное увеличение репродуктивного потенциала по сравнению с обоеполыми насекомыми сходного размера (Дьяконов, 2003). Сосущий способ питания весьма экономичен и позволяет значительную часть ассимилируемой энергии тратить на размножение. Благодаря указанным особенностям тля же быстро размножается и сильно повреждает посадки. Исходя из этих соображений, можно было бы порекомендовать более ранние сроки обрезки (например, конец апреля – начало мая), что позволит взрос-лым жукам отложить яйца на вновь отросшие молодые побеги. Подобные сроки рекомендуют и агротехники: для интенсификации отрастания побегов шиповника обрезку лучше осуществлять в период относительного покоя растения – для средней полосы России это осень (сентябрь-октябрь) или весна (март-апрель) (Серебрякова и др., 1977).
Отметим, что не охвачены плановыми работами внутриквартальные насаждения, где формовочная обрезка проводится нерегулярно либо вовсе отсутствует. Гетеротрофное население внутриквартальных интактных на-саждений шиповника развивалось в обычном режиме, для энотомофага – с массовым выходом личинок в середине июня.
Имеет значение время проведения агротехнических мероприятий, кото-рые у бюджетной организацией (работы по уходу за зелеными насаждениями в городе осуществляет организация МУП «Горзеленхоз») зависят от финансирования, и осуществляются в разные сроки в течение вегетативного сезона по мере поступления средств. Как следствие – неодновременность проведения работ в течение сезона, в разные годы, в разных частях города с оставлением нетронутых островок внутриквартальных зеленых насаждений. Получается, что агротехнические «нарушения» как бы перемещаются в пространстве и времени (по аналогии с концепцией гэп-мозаики – Коротков, 1991). Со временем отрастающие после обрезки молодые побеги восстанавливают нарушенные местообитания, делая их вновь пригодными к заселению животными – фитофагами, а следом и энтомофагами. Такая неоднородность позволяет подвижным насекомым (в том числе и тлям, и коровкам) перемещаться между участками разной стадии восстановления (с использованием внутриквартальных интактных наса-ждений в качестве рефугиев), делая возможным существование своеобразных микросукцессий заселения (иммиграции) и исчезновения («вымирания») растений – местообитаний фитофагами и энтомофагами.
Выводы:
Формовочная обрезка является эффективным приемом регуляции чис-ленности энтомофитофага (тли), но одновременно отрицательно влияет на все преимагинальные стадии развития энтомофага (кокцинеллид).
Более ранние сроки обрезки кустарника (например, конец апреля) позволит смягчить отрицательное воздействие на энтомофагов.
При выборе целей: поддержание высокой численности энтомофагов либо обеспечение эстетики и «здоровья» зеленых насаждений – преимущество следует отдать последней.
Динамика агротехнических «нарушений» в пространстве и времени обуславливает существование микросукцессий заселения растений активно перемещающимися фитофагами и энтомофагами.
СПИСОК ЛИТЕРАТУРЫ
Баранник А.П., 1979. Эколого-фаунистическая характеристика дендрофильной энтомофауны зеленых насаждений промышленных городов Кемеровской области // Экология. № 1. С. 76-79.
Дьяконов К.П., 2003. Трофические связи тлей (Homoptera, Aphidinea) как пример оптимального использования насекомыми кормовых ресурсов. // Чтения памяти А.И. Куренцова. Вып. 13. С. 53-60.
Едренкина В.А., 2005. Флора и растительность зеленой зоны города Уфы: влияние человека и задачи охраны: Дис. … канд. биол. наук. Уфа: Башкирский гос. ун-т. 317 с.
Зей-Нечаева А.Н., 1984. Энтомофауна шиповника в искусственных биогеоценозах г.Уфы // Фауна и экология животных УАССР и прилежащих районов: сб. науч. тр. / Ред.-сост. М.А.Стратийчук. Ижевск. С.103-108.
Киселев С.В., 2005. Экологические аспекты энтомофауны промышленных зон г. Тулы: Дис. … канд. биол. наук. Тула: Тульский гос. пед. ун-т. 178 с.
Колесников С.А., Болдырев М.И., 2007. Видовой состав фитофагов шиповника // АГРО XXI. № 7–9. С. 13-14.
Коротков В.Н., 1991. Новая парадигма в лесной экологии // Биологические науки. №7. С. 7–19
Кузьминов Д.В., 2005. Функциональная роль и экологическая значимость дендрофильной афидофауны в городской и пригородной зоне Воронежа. Дис. … канд. биол. наук. Воронеж: Воронежская гос. лесотехническая академия. 159 с.
Мартемьянов В.В., Бахвалов С.А., 2007. Экологические взаимосвязи в системе триотрофа и их влияние на развитие и популяционную динамику лесных филлофагов // Евразиатский энтомологический журнал. Т. 6. С. 205–221.
Савойская Г.И., 1983. Личинки кокцинеллид (Coleoptera, Coccinellidae) фауны СССР. Л.: Наука. 244 с.
Серебрякова Н.В., Орлова В.Б., Трофимец Н.В., 1977. Влияние агротехни-ческих приемов на повышение урожая плодов шиповника и улучшение их качества // Витаминные растительные ресурсы и их использование: сб. статей / Под ред. Н.Е.Зубцовского и др. М.: Наука. С. 202-210.
Тарасова О.В., 2004. Насекомые – филлофаги зеленых насаждений городов: особенности структуры энтомокомплексов, динамики численности популяций и взаимодействия с кормовыми растениями. Дис. … д-ра с.-х. наук. Красноярск: Сибирский гос. технологический ун-т, 360 c.
Ченикалова Е.В., Мохрин А.А., Щербакова С.А., 2008. Триотроф: «Зерновые–тля–кокцинеллиды» // Защита и карантин растений. №3. С. 61.
Щербакова Л.Н., Карпун Н.Н., 2008. Защита растений. Учебное пособие. – М.: Академия. 272 с.
Яблоков-Хнзорян С.М., 1976. Введение в изучение фауны кокцинеллид СССР (Coleoptera, Coccinellidae) // Зоол. сб. АН АрмССР. Т. 17. С. 101-172.
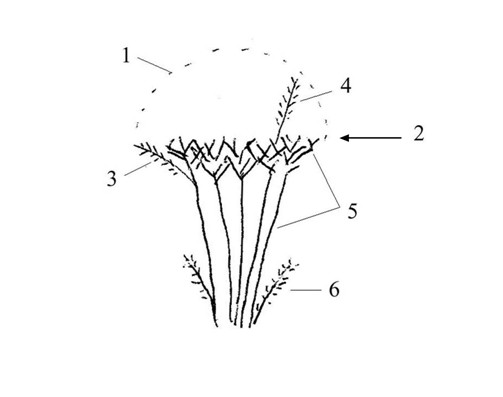
Рис. Силуэт шиповника при формовочной обрезке. 1 – область отрост-ков до обрезки; 2 – уровень плоскостной формовочной обрезки; 3 – «боковые» побеги; 4 – оставшиеся необрезанными «случайные» побеги; 5 – «скелетные» ветви; 6 – «порослевые» побеги.
MAN AS THE REGULATOR OF “PLANT – ENTOMOPHYTOPHAGE – ENTOMOPHAGE” SYSTEM IN THE URBAN ENVIRONMENT
2011 A. F. Khabibullin1, V. F. Khabibullin2
1Bashkir State Pedagogical University, Ufa 450000, Russia
2 Bashkir State University, Ufa 450074, Russia
e-mail: herpetology@mail.ru
In urban ecosystem humans by means of the agrotechnical measures, in particular, by molding trimming, influence the sanitary and decorative state of urban green plantings. The ambiguity of human activity in management for each members of triotroph system “plant – entomophytophage – entomophage” is shown. The agrotechnical measures are positive for both man and plant, but negative for phytophagous and entomophagous insects. Dynamics of agrotechnical “disturbances” in space and time can cause the existence of micro successions of plant occupation by insects.
Key words: urban ecosystem, triotroph “plant – entomophytophage – entomophage”, urban green plantings, Rosa sp., Macrosiphum rosae, Adalia bipunctata.
Добавить комментарий