Хабибуллин В.Ф., Хабибуллин А.Ф.
Зоол. журн. 2011. Т.90. №10. С.1180-1187
Обсуждается «механический» аспект рекреационного воздействия, под которым понимается физическое контактное воздействие антропогенной природы, оказываемое на биоту. Исследовано влияние механического аспекта фактора беспокойства на оборонительное поведение насекомых-хортобионтов на примере двух видов кокцинеллид с разной экологией. Сделаны выводы о существовании характерных тактических приемов оборонительного поведения, набор и специфика применения которых зависят от видовой принадлежности. Выявлены различные оборонительные стратегии: хищные виды применяют деятельную стратегию (легкость падения и быстрота возобновления активности), грибоядные – инертную стратегию («неохотность» падения и медленное возобновление активности, зачастую с танатозом). При механическом воздействии адаптивными могут считаться поведенческие реакции, позволяющие минимизировать траты времени и энергии при поддержании приемлемого уровня безопасности. В условиях рекреации при ослаблении действия ограничивающих факторов на первый план выходит рациональность бюджета энергии и времени.
Ключевые слова: рекреация, механический аспект, оборонительное поведение, хортобионты, Coccinellidae, Psyllobora vigintiduopunctata, Coccinula quatuordecimpustulata.
Антропогенное воздействие на естественные экосистемы весьма разнообразно, поэтому вполне закономерно рассмотрение различных ее сторон (Гашев, 2000), в частности, рекреационного воздействия. Содержание понятия «рекреация» варьирует в различных отраслях знания и в зависимости от ракурса рассмотрения. Мы возьмем за основу следующее рабочее определение (Сионова, 2005): «Рекреация – воздействие человека на окружающую среду во время отдыха». Рекреационное воздействие может проявляться в вытаптывании травы, повреждении древесно-кустарниковой растительности, нарушении мест гнездовий и укрытий животных, шумовом загрязнении, вносе и выносе вещества и энергии и др. Воздействие рекреации на животных может быть как прямым, или непосредственным (уничтожение, преследование, беспокойство), так и непрямым, или опосредственным, и осуществляться через среду обитания (растительность, почву, гидрологический режим, пищевые ресурсы) (Cole, Landres, 1995). На наш взгляд, принципиально важна трактовка понятия рекреации именно как деятельности, в первую очередь, физической активности человека на лоне природы (см. Саранча, 2009: «Рекреация, прежде всего, представляет собой деятельность субъекта»). Рекреация как таковая может служить объектом и целью исследований, но также может выступать как удобная модель для изучения ряда общетеоретических проблем и более частных вопросов.
В области изучения антропогенных воздействий преобладают работы токсикологической направленности (Нестерков, 2009); одним же из наименее изученных аспектов антропогенного воздействия вообще и рекреационного в частности является так называемый «механический». Под данным термином мы понимаем физическое контактное воздействие антропогенного характера, оказываемое на компоненты живой природы. Под это определение подпадает преимущественно прямое воздействие человека и техники, а также, если смотреть шире, любая активность человека, которая механически воздействует на биоту.
Механический аспект антропогенного воздействия (в сублетальных дозах) можно трактовать через категорию «фактор беспокойства». Виды фактора беспокойства можно выделять по различным основаниям: по природе он может быть механическим, химическим, акустическим, оптическим и пр.; на субъектной основе можно выделять многообразие агентов – элементарных раздражителей: человек, домашние животные, машины и механизмы; по длительности воздействия – постоянно, временно, кратковременно действующие факторы беспокойства (Давыгора, 2000). Весьма полезным представляется конкретизация этого понятия Владышевским (2004): «Фактор беспокойства – воздействие на животных внешних раздражителей, воспринимаемых в качестве сигнала опасности». Для беспозвоночных (в частности, свободноживущих насекомых), на наш взгляд, вполне применима известная формула бихевиористов «стимул-реакция» (Гороховская, 2001): насекомые реагируют не на фактор беспокойства (стрессор) вообще, но на конкретные стимулы, обусловленные этим стрессором; и при этом нет принципиальных различий между природными и антропогенными стимулами.
Объектами при изучении влияния рекреации на биоту выступают, как правило, позвоночные животные (Захаров, 1998; Жигарев, 2005 и др.), беспозвоночные в этом плане изучены слабее. Вопросы механического воздействия травоядных млекопитающих, равно как человека, машин и механизмов на хортобионтов остаются не изученными.
Считается, что при рекреации в первую очередь страдает травяной ярус и обитающие в нем животные – хортобионты и хортофилы (по терминологии Лагунова, 2008). Ведущим прямым фактором при этом служат механическое воздействие, которое отдыхающие как агенты фактора беспокойства оказывают на хортобионтов.
Животные как организмы, обладающие поведением, способны реагировать на внешние воздействия стрессоров тремя принципиальными путями (Биоиндикация …, 1988): пространственное и (или) временное избегание; использование конституциональных особенностей тела, включая специ-фическую двигательную активность; изменение свойств окружающей среды. Первый и тем более третий путь нечасто встречается у беспозвоночных; мы остановимся на обсуждении второго пути – двигательной активности.
В данной работе мы исследуем влияние механического аспекта фактора беспокойства на оборонительное поведение насекомых-хортобионтов.
Первой нашей задачей было выявление элементов (тактических приемов) оборонительного поведения, демонстрируемого насекомыми при воздействии механического стрессора. Вторая задача сформулирована нами так: проверить, различаются ли применяемые тактические приемы у разных видов (иными словами, существуют ли в разных стратегиях оборонительного поведения). Третья задача: выяснить, насколько эффективны выделенные поведенческие стратегии в условиях механического рекреационного воздействия.
МАТЕРИАЛ И МЕТОДЫ
Материал собран 19-20 июля 2009 г. на территории одной из баз отдыха, расположенных в Южном Предуралье (Республика Башкортостан) близ пос. Зирган, на левом берегу р. Белой (53º16’ с.ш. 55 º19’ в.д.).
Объект исследования – жуки сем. кокцинеллид (Coleoptera; Coccinellidae) двух видов: двадцатидвухточечная Psyllobora vigintiduopunctata (L. 1758) и четырнадцатипятенная Coccinula quatuordecimpustulata (L. 1758) божьи коровки. Далее для краткости будем называть их соответственно псиллобора и кокцинула. Оба вида – типичные хортобионты; обычны в районе исследования, имеют средние размеры (3.5 – 4 мм), активно не летают во время поиска пищи, хорошо заметны. Различия заключаются в трофической специализации: первый вид – мицетофаг (объект питания – мучнистая роса), второй – энтомофаг (объект питания – тля). Хищные кокцинулы гораздо подвижнее, активно перемещаются в поисках пищи, а грибоядные псиллоборы малоподвижны. Объем материала: 22 особи кокцинул и 54 – псиллобор.
Работа проведена на опушке лиственного редколесья, на разнотравном участке общей площадью около 100 м2 (более освещенная часть – с вкраплениями злаковых – местообитание кокцинул, менее освещенная, с пятнами светотени – местообитание псиллобор). Температура воздуха в период работ 22-25ºС, ветер слабый, облачность небольшая.
В качестве механического агента воздействия мы рассматривали человека, который при хождении по траве оказывает физическое (механическое) воздействие на насекомых – хортобионтов. Имитация такого воздействия экспериментально осуществлялась следующим образом: после визуального выявления насекомого по этому участку травостоя однотипно проводили рукой (не резко, со скоростью как при спокойной ходьбе), механически воздействуя на насекомое. Задевается ли вскользь само насекомое – дело случая и зависит от расположения жука на растении относительно вектора воздействия. Если насекомое не падало после первого акта механического воздействия, его повторяли до момента падения (это позволяет судить о том, насколько крепко насекомое удерживается на растении). Для грубой оценки риска механического повреждения упавших с травы на землю жуков мы наступали на 15 случайным образом выбранных кокцинеллид: как находящихся в состоянии танатоза, так и подвижных (исходили из основанной на предварительных наблюдениях предположении о безвредности такой манипуляции для жуков в условиях травостоя).
Мы экспериментально фиксировали следующие реакции жуков на механическое беспокойство.
1. Замечают ли кокцинеллиды приближающегося человека и улетают ли при его приближении?
2. Удерживается ли насекомое на растении при механическом воздействии?
3. Во время падения пытается ли расправить крылья и улететь, или пытается зацепиться за части растения?
4. Впадает ли в состояние танатоза? Через какое время после падения возобновляет активность?
Измеряли время с момента падения на землю до момента возобновления активности (с точностью до 1 с). Каждую особь тестировали только один раз (если специально не оговорено). Копулирующие пары анализировали совместно с одиночными.
Отмечали наличие или отсутствие состояние танатоза. Танатоз (по Реймерсу, 1988) – это защитная поведенческая реакция животных, имитация отсутствия жизни. Кратковременное состояние неподвижности насекомых – небольшой временной лаг между падением и возобновлением нормальной активности – мы трактовали как «затаивание». В литературе мы не обнаружили четких критериев – когда заканчивается затаивание и начинается танатоз, длительность затаивания изменяется постепенно, и объективный выбор порогового значения затруднен. Мы взяли в качестве порогового значение в 5 с: неподвижное состояние длительностью до 5 с включительно трактовали как затаивание, свыше 5 с – как танатоз.
Статистическая обработка данных осуществлена с использование пакета программ MS Excel 2003 и Statistica 5.1. Для оценки достоверности различий использовали непараметрические критерии: χ-квадрат, U-критерий Манна-Уитни.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Насекомые существуют в специфическом мезомире и реагируют на воздействия агентов фактора беспокойства в пределах относительно небольшого радиуса восприятия стимулов (визуальных, акустических, химических, вибрационных, механических и др.). Для выяснения вопроса, реагируют ли кокцинеллиды на приближающегося человека, мы сначала рассмотрим, какие именно стимулы они способны дистантно воспринимать, а также их основные поведенческие реакции на эти стимулы.
Оптические стимулы. У кокцинеллид обнаружены визуальные рецепторы трех типов (Lin, 1993), чувствительных к синим (420 нм), зеленым (520 нм) и ультрафиолетовым (360нм) волнам с общим диапазоном 350-700 нм и максимумом при 500-600 нм. Характерно, что радиус обнаружения пищевого объекта невелик: 0.7 см для личинок и около 1 см для имаго (Stubbs, 1979). Коровки способны дистанционно распознавать контур крупных предметов: так, их привлекает силуэт дерева (Hattingh, Samways, 1995). Из собственного опыта наблюдений мы можем заключить, что крупные виды кокцинеллид (более 4 мм в длину, например, семиточечная или глазчатая (Anatis ocellata (L. 1758)) коровки) визуально замечают человека и реагируют на его приближение на расстоянии 0.4-0.6 м; средние и тем более мелкие виды (до 4 мм в длину – из родов Stethorus, Scymnus s. lato, Coccinula, Psyllobora, Tytthaspis) визуально не реагируют на человека на расстоянии 0.25-0.4 м.
Ольфакторные (химические) стимулы. Экспериментально показана реакция кокцинеллид-афидофагов на запах кормового растения (на котором питаются пищевые объекты коровок – тли), которое, вероятно, испускает аттрактанты, привлекающие коровок (Schmid, 1992). Выявлена реакция и на экстракт кормовых растений, а также запах самих пищевых объектов – тлей (Schaller, Nentwig, 2000).
В доступной литературе мы не обнаружили сведений о восприятии кокцинеллидами акустических стимулов. По нашим наблюдениям, кокцинеллиды не реагируют на шумы фоновой активности человека – разговор, смех; при отсутствии резких запахов парфюма химическим воздействием человека также можно пренебречь. Так, прямо перед центральным входом главного комплекса базы отдыха (место сбора материала – см. раздел «Материал и методы») растет сосна обыкновенная (Pinus silvestris L. 1753), на которой многочисленны все стадии развития десятиточечной (Adalia decimpunctata (L. 1758)) и двуточечной (Adalia bipunctata (L. 1758)) коровок. Выраженное антропогенное воздействие – с утра до 2-3 ч. ночи: постоянные разговоры, крики, смех, плач детей, ругань, музыка, шум, свист, ночное освещение, курение в двух метрах от ствола (т.е. нижние ветви окуриваются) – не приводят к снижению численности и исчезновению коровок.
Механические стимулы. Как правило, при слабых тактильных воздействиях кокцинеллиды принимают пассивно-оборонительную позу, подбирая под себя конечности и плотно прижимаясь телом к субстрату. При сильных тактильных воздействиях кокцинеллиды впадают в состояние оцепенения, или танатоза.
На основании наблюдений в моделируемой ситуации механического воздействия мы выделили следующие тактики оборонительного поведения (как выбор альтернативных вариантов поведения).
1. При возможности дистантного обнаружения приближающегося агента беспокойства у насекомого есть альтернатива: улететь или остаться.
2. При непосредственном механическом воздействии (момент контакта) есть альтернатива: удержаться на растении или начать падение на землю
3. Во время падения есть альтернатива: расправить крылья и улететь или продолжить падение до земли.
4. После падения на землю есть альтернатива: замереть/впасть в состояние танатоза или начать сразу же активное движение.
Обсудим применяемые модельными видами тактические приемы.
Тактика № 1. У модельных видов эта тактика не выявлена: не было отмечено ни одной попытки улететь при приближении человека (что и неудивительно, учитывая «близорукость» коровок – см. выше). Подобная тактика в основном характерна для видов пустынь (Савойская, 1983).
Тактика № 2. Подавляющее число кокцинеллид (в сумме 97.4% по обоим видам), в том числе 100% кокцинул и 90.9% особей псиллобор (различия достоверны при p<0.05) падали после первого же механического воздействия. Две особи псиллоборы крепко удерживались на растении и упали лишь после четвертого и пятого акта воздействия.
Тактика № 3. Взлет во время падения отмечен только два раза (в обоих случаях это были псиллоборы, которые в дальнейших расчетах не учитывались). Однако необходимо отметить, что оба раза это происходило на единичных высоких (более 30 см) растениях (средняя высота травостоя в месте выполнения работ около 12-20 см). Для выяснения этого вопроса необходимо тестировать всю выборку жуков на высоких растениях, что не было возможности осуществить в наших условиях. Попутно добавим, что мелкие виды кокцинеллид (из рода Scymnus s. lato) регулярно применяют эту тактику – взлет во время падения.
Тактика № 4. Псиллоборы несколько чаще впадают в состояние танатоза, чем кокцинулы (38.5% против 31.8%; различия статистически не значимы).
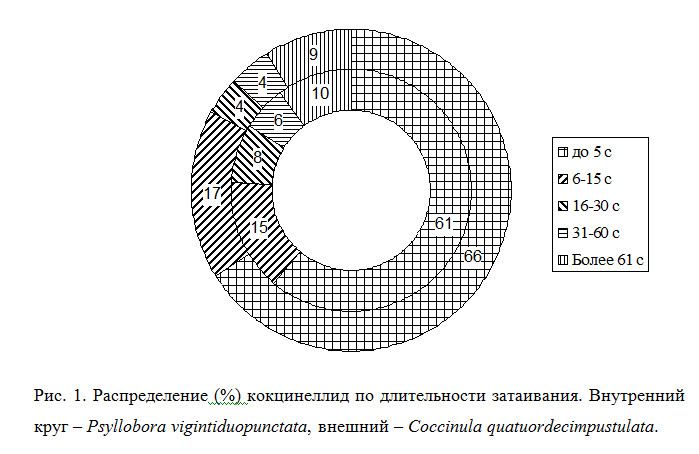
Значения скорости возобновления активности после падения следующие (рис. 1). У псиллобор (n=52) среднее время возобновления активности – через 15.5 с после падения (lim 0-121 с); у кокцинул (n=22) среднее время возобновления активности через 11.2 с (lim 0-103 с) (различия недостоверны). Если же сравнивать отдельно особей, не впавших в состояние танатоза (т.е. возобновивших свою активность в первые пять секунд после падения на землю), то выясняется, что кокцинулы быстрее возобновляют активность: в среднем через 0.8 с против 1.8 с (различия статистически достоверны при p<0.05). Показательно и то, что доля кокцинул, возобновивших активность сразу же после падения (временной лаг 0 секунд) достоверно выше, чем у псиллобор (11 особей, или 50% против 8 особей, или 15.4%, p<0.01).
Ни одна из протестированных копулирующих пар (1 пара кокцинул и 8 пар псиллобор) не распались при механическом воздействии. После падения самка кокцинулы возобновила активность через 2 с, самки псиллобор в среднем через 40 с (lim 0-102 с). Самцы in copuli остаются неподвижны; лишь в одном случае самец псиллоборы начал проявлять активность, перебирая передними конечностями, раньше самки (через 62 против 102 с у самки).
Любая поведенческая стратегия, в том числе оборонительная, реализуется через тактику – посредством набора конкретных приемов и действий. Защитные механизмы насекомых разнообразны (см., например, сайт «Жуки (Coleoptera) и колеоптерологи»); одна из самых известных – это наличие предостерегающей окраски. Однако при механическом воздействии на насекомых их апосематическая окраска не выполняет своей защитной функции: как естественные агенты (проходящие мимо крупные дикие животные, ветер, дождь), так и антропогенные агенты (домашние животные, машины и механизмы, сам человек) воздействуют на насекомых независимо от наличия или отсутствия у них предостерегающей окраски. В данном случае важное значение приобретают поведенческие механизмы.
Основываясь на интерпретации полученных данных по тактическим приемам, у модельных видов кокцинеллид мы выделили две различные оборонительные стратегии:
Первая (условно пассивная) – поведение насекомых характеризуется следующими чертами:
– при механическом воздействии крепче удерживаются на растении,
– большее время находятся в состоянии оцепенения (т. е. медленнее возобновляют активность),
– чаще впадают в состояние танатоза.
Вторая (условно активная) – поведение насекомых характеризуется следующими чертами:
– при механическом воздействии слабее удерживаются на растении – падают на землю даже при незначительном по силе механическом воздействии,
– меньшее время находятся в состоянии оцепенения (т. е. возобновляют активность практически сразу после падения),
– реже впадают в состояние танатоза.
Некоторые затруднения возникли при именовании выделенных стратегий: термины «активная» и «пассивная» – слишком широко употребительные, их содержание размыто. Поэтому в качестве уточняющих мы предлагаем использовать термины «деятельная» для условно-активной и «инертная» для условно-пассивной стратегий, которые не так часто используется в биологических науках, их значение не столь расплывчато и может быть конкретизировано.
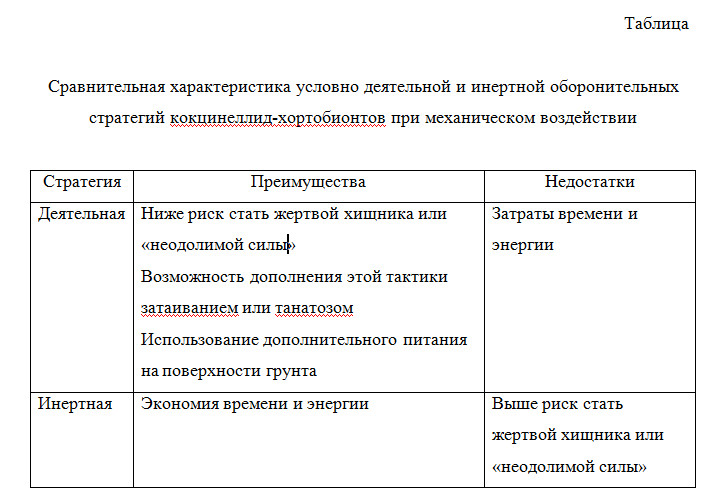
Формирование стратегии оборонительного поведения животных в ответ на действие стрессорных стимулов происходит под влиянием двух противоположных мотивов: с одной стороны, это стремление обеспечить собственную безопасность, с другой – стремление минимизировать потери времени и энергии, затрачиваемые на достижение этой безопасности (см., например, Владышевский, 2004). Эти противоречия преломляются в преимущества и недостатки (таблица), свойственные стратегиям с уклоном в сторону максимизации безопасности или минимизации потерь времени и энергии.
Удерживаясь на растении, насекомые не тратят энергии на поиск подходящего растения, взбирание на него и поиск подходящего места на растении. Так как для насекомых энергетически невыгодно при каждом порыве ветра падать на землю и затем тратить силы и время на карабкание, то, по-видимому, отбор идет на закрепление прочности удерживания на растении. С другой стороны, удерживаясь на растении, насекомые увеличивают риск пострадать от агента механического воздействия – например, если это энтомофаг (риск быть съеденным), крупные травоядные (риск быть случайно проглоченным вместе с травой) или просто «неодолимая сила». Возможно, при сильных порывах ветра (видимо, существует некое пороговое значение интенсивности колебаний), а также при прямом физическом контакте адаптивнее не продолжать удерживаться на растении, а упасть на землю.
Падая на землю, насекомое существенно снижает риск стать жертвой хищника или «неодолимой силы» (риск быть раздавленным или покалеченным крупным массивным объектом – позвоночным животным, человеком, механизмом). В ряде случаев (при воздействии неодолимой силы) этого бывает достаточно, но чтобы наверняка обезопасить себя от возможного нападения хищника, можно дополнить эту тактику затаиванием или танатозом. Интересно, что для кокцинеллид у падения есть и другие плюсы. Так как механическое воздействие одинаково для всех хортобионтов, то некоторые тли также используют тактику падения, и могут послужить добычей для упавших на землю афидофагов. Так, в пшеничных полях 30% жертв семиточечной коровки – тли, передвигающиеся по земле (Ferran, Dixon, 1993).
Выбор стратегии зависит, на наш взгляд, от экологии вида (прежде всего трофической): кокцинулы как активные хищники применяют деятельную стратегию, а псиллоборы как малоподвижные, питающиеся прикрепленными пищевыми объектами насекомые, применяют инертную стратегию.
В чем состоит специфика механического воздействия на хортобионтов в условиях рекреации?
Во-первых, она проявляется на общем рекреационном фоне, т.е. имеет место наличие всех атрибутов рекреации: беспокойство, вытаптывание, замусоривание. Беспокойство проявляется прежде всего в механическом воздействии, влиянием замусоривания при надлежащей организации отдыха можно пренебречь; интенсивность вытаптывания условимся считать низкой, не приводящей к деградации фитоценозов (опосредственное растительностью и почвой влияние рекреации на животное население мы здесь не рассматриваем). По-видимому, преувеличивается и физическое влияние отдыхающих на хортобионтов: подвижные группы насекомых (прыгающие, летающие) активно избегают физического контакта, малоподвижные затаиваются или скрываются в подстилке. Прямое физическое давление также не является значимым фактором. По разным оценкам, статическое давление человека на грунт составляет 180-375 г/см2 (Liddle, 1997), причем на траве эти показатели снижаются. Нами при изучении давления, оказываемого телом человека на находящихся на земле среди травы насекомых, внешних повреждений, а также изменений в последующей активности не выявлено ни у одной из 15 протестированных особей кокцинеллид. Наблюдения за особями непосредственно после экспериментальных воздействий видимых отрицательных для жуков последствий не выявили: все особи кокцинеллид продолжали рутинную активность. Изменений в плотности населения коровок на модельном участке до и после проведения работ не зафиксировано. Отдельно должен рассматриваться вопрос об интенсивности рекреационного воздействия на природные экосистемы (что не входит в круг обсуждаемых нами задач); мы лишь заметим, что в местах организованного отдыха с продуманной дорожно-тропиночной сетью номинальные показатели интенсивности рекреационной нагрузки (например, 40 чел/га) именно для травостоя могут быть значительно (в 1.5-2 раза) выше реальных.
Во-вторых, с точки зрения безопасности хортобионтов важны следующие обусловленные организованной рекреацией факторы:
– ограничение движения и работы машин и механизмов;
– отсутствие крупных травоядных млекопитающих – как диких, так и домашних;
– снижение пресса насекомоядных позвоночных (в литературе обычно отмечается снижение разнообразия и численности позвоночных в целом и насекомоядных в частности (Сионова, 2005 и др.)).
Эти факторы (назовем их ограничивающие) напрямую влияют на безопасность и выживание хортобионтов, причем первые два – исключительно механически. Рекреация, уменьшая или полностью нивелируя их действие, снижает давление отбора в направлении достижения максимальной безопасности.
Помимо модификации имеющихся, рекреация привносит собственный «механический» фактор, влияющий на эффективность поведенческих оборонительных стратегий: человек, двигаясь по траве, неизбирательно физически воздействует на всех оказавшихся на его пути хортобионтов; однако целенаправленно никого не преследует хортобионтов и не уничтожает. Этот фактор не приводит к элиминации хортобионтов, при этом влияет на бюджет времени и энергии.
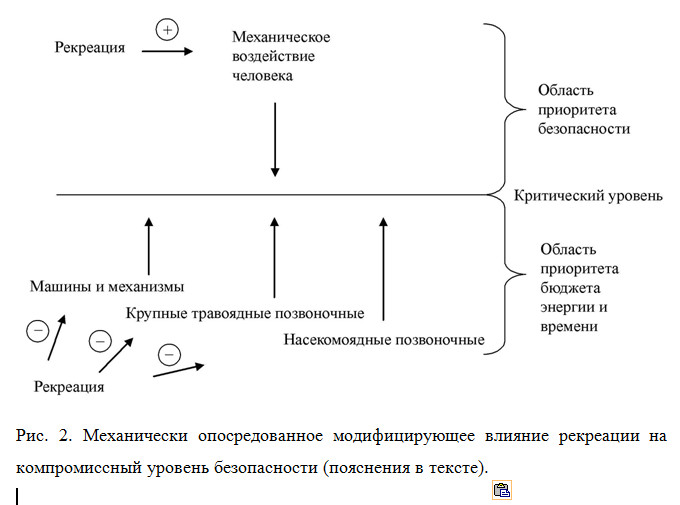
Принципиальная схема такова (рис. 2): существует «критический уровень» –компромисс между максимизацией безопасности животного и минимизацией затрат энергии и времени на обеспечение безопасности, на достижение и поддержание которого направлено оборонительное поведение хортобионтов. С одной стороны, рекреация ослабляет (на схеме отмечено знаками «минус») давление естественных векторов отбора (в нашем случае материализованных в виде машин и механизмов, крупных травоядных и насекомоядных позвоночных), существенных для безопасности насекомых-хортобионтов. С другой стороны, рекреация добавляет (знак «плюс») новый вектор отбора в виде индифферентного механического воздействия человека, который не является критическим для безопасности, но влияет на бюджет времени и энергии. Ослабляя действие естественных факторов, он смещает границу компромиссного уровня безопасности из зоны приоритета безопасности в зону приоритета рационального бюджета энергии и времени. Таким образом, рекреация опосредованно через контрбаланс упомянутых выше векторов «уравновешивает» оборонительное поведение хортобионтов в области некоего компромиссного «критического уровня».
Итак, в условиях механического воздействия адаптивными могут считаться поведенческие реакции, позволяющие минимизировать траты времени и энергии при поддержании приемлемого уровня безопасности. В нашем случае этими реакциями можно считать: удерживание на растении (ибо падение – это трата и времени, и энергии), в случае же падения на землю – немедленное возобновление активности (минимизация потерь времени). Первая реакция характерна для инертной стратегии, вторая – для деятельной.
Несколько предпочтительней кажется инертная стратегия, которая в обсуждаемых условиях практически лишена указанных нами выше в таблице недостатков, т.к. меньше источников риска: в местах организованного отдыха меньше пресс насекомоядных позвоночных; нет крупных травоядных; ограничено движение машин и механизмов. Преимущества же деятельной стратегии в обсуждаемом ракурсе (см. таблицу) работают вхолостую: меньшее число насекомоядных позвоночных и меньшая вероятность воздействия «неодолимой силы». Правда, «прочность удержания» на растении у модельных видов незначительна (3.7%), соответственно, и ее актуальная роль невелика.
С другой стороны, адаптации, направленные на минимизацию потерь энергии, увеличивают потерю времени или снижают безопасность (или наоборот). Подобный конфликт интересов или, в терминологии эволюционной теории, контрбаланс векторов отбора (Северцов, 1998) отражает противоречивость отдельных поведенческих оборонительных реакций насекомых-хортобионтов и сложность их объединения в рамках единой стратегии.
Данная схема, изложенная исходя из общих дедуктивных рассуждений, в дальнейшем может быть верифицирована и скорректирована частными эмпирическими экспериментами. В перспективе можно попытаться количественно оценить интенсивность факторов, например, в системе координат (x; y; z): (уровень безопасности; уровень энергетических затрат; потери времени), придавая каждому измерению различные относительные веса.
Выводы:
1. У насекомых-хортобионтов существует набор тактических приемов оборонительного поведения, набор и специфика применения которых зависят от видовой принадлежности.
2. Выявлены различные оборонительные стратегии: хищные кокцинулы применяют деятельную стратегию: легкость падения и быстрота возобновления активности, грибоядные псиллоборы – инертную стратегию: «неохотность» падения и медленное возобновление активности, зачастую с танатозом.
3. В условиях умеренной рекреации при ослаблении действия ограничивающих факторов на первый план выходят не вопросы безопасности, а бюджет энергии и времени; именно рациональное использование времени является преимуществом деятельной стратегии.
СПИСОК ЛИТЕРАТУРЫ
Биоиндикация загрязнений наземных экосистем, 1988. Пер. с нем. М.: Мир. 350 с.
Владышевский А.Д., 2004. Значение фактора беспокойства для диких птиц и млекопитающих. Дис. … канд. биол. наук. Красноярск: Красноярский гос. ун-т. 230 с.
Гашев С.Н., 2000. Млекопитающие в системе экологического мониторинга (на примере Тюменской области). Тюмень: Изд-во Тюменского гос. ун-та. 220 с.
Гороховская Е.А., 2001. Этология: рождение научной дисциплины. СПб.: Алетейя. 224 с.
Давыгора А.В., 2000. Фактор беспокойства и характер его воздействия на степного орла в Предуралье // Труды института биоресурсов и прикладной экологии. Вып. 1. С. 107-134.
Жигарев И.А., 2005. Организация и устойчивость рекреационных сообществ: на примере мелких млекопитающих. Дис … док. биол. наук. М.: Московский государственный педагогический университет. 455 с.
Захаров В.Д., 1998. Влияние рекреационной нагрузки на сообщества птиц в лесах Южного Урала // Известия Челябинского научного центра УрО РАН. № 1. С. 141-150.
Лагунов А.В., 2008. Стратиграфия хортобионтных полужесткокрылых Ильменского заповедника (Южный Урал) // Принципы и способы сохранения биоразнообразия: Материалы III Всерос. науч. конф. Йошкар-Ола. С. 67-68.
Нестерков А.В., 2009. Реакция населения беспозвоночных-хортобионтов луговых сообществ Среднего Урала на выбросы медеплавильного производства. Автореферат дис. … канд. биол. наук. Екатеринбург: Ин-т экологии растений и животных УрО РАН. 20 с.
Реймерс Н.Ф., 1988. Основные биологические понятия и термины. М.: Просвещение. 319 с.
Савойская Г.И., 1983. Личинки кокцинеллид фауны СССР. Л.: Наука. 244 с.
Саранча М.А., 2009. Проблема концептуального определения и соотношения понятий «туризм» и «рекреация» // Вестник Удмуртского ун-та. Сер. Биология. Науки о земле. Вып. 2. С. 105-118.
Северцов А.С., 1998. Эволюция популяций и эволюция биоценозов // Зоол. журн. Т. 77. № 5. С. 517-526.
Сионова М.Н., 2005. Влияние рекреации на биоразнообразие модельных групп организмов нижнего яруса широколиственных и сосновых лесов Калужской области. Дис. … канд. биол. наук. Калуга: Калужский государственный педагогический университет. 151 с.
Cole D.N., Landres P.B., 1995. Indirect Effects of Recreation on Wildlife // Wildlife and Recreationists – Coexistence Through Management and Research. Washington, DC: Island Press: Chapter 11. P. 183-202.
Ferran A., Dixon A.F.G., 1993. Foraging behaviour of ladybird larvae (Coleoptera; Coccinellidae) // Eur. J. Entomol. V. 90. P. 383-402.
Hattingh V., Samways M.J., 1995. Visual and olfactory location of biotopes, prey patches, and individual prey by the ladybeetle Chilocorus nigritus // Entomol. Exp. Appl. V. 75. P. 87-98.
Liddle M., 1997. Recreation ecology: the ecological impact of outdoor recreation and ecotourism. London, Chapman & Hall. 639 p.
Lin J.-T., 1993. Identification of photoreceptor locations in the compound eye of Coccinella septempunctata Linnaeus (Coleoptera, Coccinellidae) // Journal of Insect Physiology V. 39 (7). P. 555-562.
Schaller M., Nentwig W., 2000. Olfactory orientation of the seven-spot ladybird beetle, Coccinella septempunctata (Coleoptera: Coccinellidae): Attraction of adults to plants and conspecific females // Eur. J. Entomol. V. 97. P. 155-159.
Schmid A., 1992. Untersuchungen zur Attraktivität von Acker¬wildkräutern für aphidophage Marienkäfer (Coleoptera, Coccinellidae) // Agrarökologie. V. 5. P. 1-122.
Stubbs M., 1979. Another look at prey detection by coccinellids // Ecological Entomology. V. 5 (2). P. 179-182.
Сайт Зоологического института РАН «Жуки (Coleoptera) и колеоптерологи» www.zin.ru/animalia/coleoptera
THE MECHANICAL ASPECT OF RECREATION AND DEFENSIVE BEHAVIORAL STRATEGIES IN GRASS-DWELLING INSECTS
2011 V. F. Khabibullin1, A. F. Khabibullin2
1 Bashkir State University, Ufa 450074, Russia
2Bashkir State Pedagogical University, Ufa 450000, Russia
e-mail: herpetology@mail.ru
Добавить комментарий